�����ڡ���ϸ���Ĵ�л
һ��Ѫ���ص�����ϳ�
�����ϸ���У�Ѫ�쵰��(hemoglolin,Hb)ռ��ϸ���ڵ�����������95%������ѪҺ����O2������Ҫ���ʣ���CO2����������һ����ϵ��Ѫ�쵰������4���ǻ���ɵ��ľ��壬ÿһ�ǻ���һ�����鵰��(globin)��һ����Ѫ����(heme)�϶��ɡ������鵰������ϳ���һ�㵰������ͬ����˱����ص����Ѫ���ص�����ϳɡ�
Ѫ����Ҳ������һЩ�����ʣ��缡�쵰��(myoglobin)����������ø(catalase)����������ø(peroxidase)�ȵĸ����������һ��ϸ�����ɺϳ�Ѫ���أ��Һϳ�ͨ·��ͬ�����˺�ϸ���У�Ѫ���صĺϳɴ��綯��ϸ����ʼ��ֱ����֯��ϸ�����Կɺϳɡ��������ϸ��������Ѫ���صĺϳɡ�
(һ)Ѫ���صĺϳ�ͨ·(����)
Ѫ���غϳɵĻ���ԭ���Ǹʰ��ᡢ��������øA��Fe�������ϳɵ���ʼ����ĩ���̾��������壬���м���ڰ�Һ�н��С��ϳɹ��̷�Ϊ�����ĸ����裺
1.��-����-��-ͪ����(��-aminplevulinicacid,ALA)�����ɣ����������У������ɸʰ������������øA��ALA�ϳ�ø(ALa synthetase)�Ĵ�����������ALA��ALA�ϳ�ø�������ǻ���ɣ�ÿ���ǻ�������Ϊ60��000���丨øΪ��������ȩ����øΪѪ���غϳɵ�����ø����Ѫ���صķ������ơ�

2.߲��ԭ�����ɣ����������ɵ�ALA�����Һ�У���ALA��ˮø(ALa dehydrase)�Ĵ��£�������ALA��ˮ���ϳ�һ����߲��ԭ(prophobilinogen,PBG)��ALA��ˮø�ɰ˸��ǻ���ɣ�������Ϊ26��Ϊ���ϻ�ø��
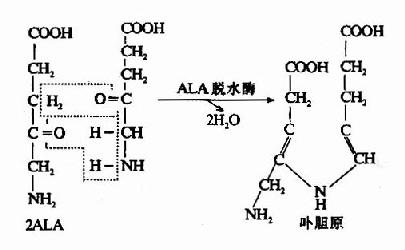
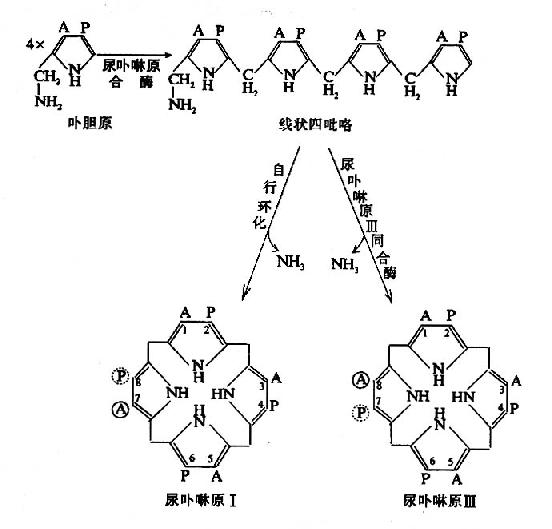
3.��߲��ԭ�ͷ�߲��ԭ�����ɣ��ڰ�Һ�У��ķ���PBG�Ѱ���������һ������߲��ԭ��(uroporphyrinogen ��UPG ��)���˷�Ӧ����������ø����߲��ԭ��ø(uroporphyrinogen synthetase)�ֳ�߲��ԭ�Ѱ�ø(PBGdeaminase)����߲��ԭ��ͬ��ø(uroporphyrinogen�� cosynthase)�����ȣ�PBG����߲��ԭ��ø�����£��Ѱ�����������״��������������߲��ԭ��ͬ��ø��������������߲��ԭ������߲��ԭ��ͬ��øʱ����״����������Ȼ��������߲��ԭ��(UPG-��)��������߲��ԭ���������ڣ�UPG���7λ��ϵ������������8λΪ���������UPg ������֮�෴����7λ�DZ��������8λ������������������UPG������UPG����Ϊ10000��1��
ʽ��A�����������P���������
��߲��ԭ���һ������߲��ԭ������ø����ʹ���ĸ������(A)���ȱ�Ϊ��(M)���Ӷ����ɷ�߲��ԭ��(coproporphyrinogen ��CPG ��)��
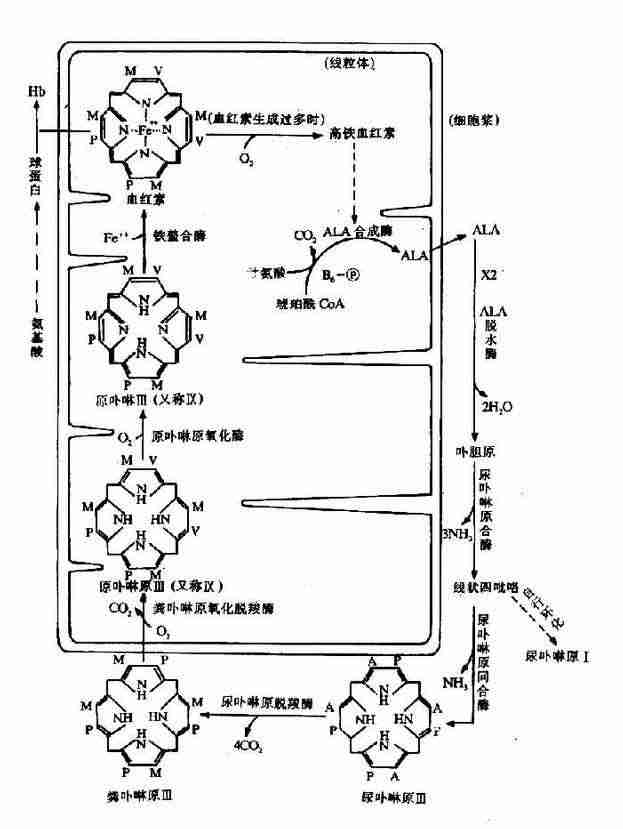
4.Ѫ���ص����ɣ���Һ�����ɵķ�߲��ԭ���ٽ����������У��ڷ�߲��ԭ��������ø�����£�ʹ2��4λ�ı����(P)��������������ϩ��(V)������ԭ߲��ԭIX���پ�ԭ߲��ԭIX����ø�����⣬ʹ����4���������ļ�ϩ�������ɼ�Ȳ��������ԭ߲��IX���������������ø(ferrochelatase)���º�Fe2���������Ѫ���ء�(ͼ10-18)��
A.�������P.�������M.����V.��ϩ��
ͼ10-18��Ѫ���ص�����ϳɼ������
Ѫ�������ɺ��������ת���Һ�����鵰��϶���ΪѪ�쵰�ס���������ÿ��ϳ�6��Hb���൱�ںϳ�210mgѪ���ء�
(��)Ѫ���غϳɵĵ���
Ѫ���صĺϳ��ܶ������صĵ��ڣ�������Ҫ�ǵ���ALA�����ɡ�
1.ALA�ϳ�ø��Ѫ���غϳ�øϵ�У�ALA�ϳ�ø������ø���������١�Ѫ���ضԴ�ø�з����������á�Ŀǰ��Ϊ��Ѫ���������ڿ����������ϣ��γ��л��Ե�������ף��Ӷ�����ALA�ϳ�ø�ĺϳɡ����⣬Ѫ���ػ�����ֱ�ӵĸ���������ALA�ϳ�ø���Ե����á�ʵ�������Ѫ����Ũ��Ϊ5��10��6Mʱ�������ALA�ϳ�ø�ĺϳɣ�Ũ��Ϊ10-5��10-4Mʱ�������ø�Ļ��ԡ����������Ѫ�������ɺ�ܿ����鵰��ϣ�����Ѫ���غϳɹ���ʱ��������Ѫ���ر�����Ϊ����Ѫ����(hematin)��������ALA�ϳ�ø��ǿ�����Ƽ������һ������ALA�ϳ�ø�ĺϳɡ�
���Լ��ء���غ��ͪ�ڸ���5��-��ԭø�����¿�����5��-��غ��ͪ�����߿��յ�ALA�ϳ�ø�IJ������Ӷ��ٽ�Ѫ���ص����ɡ�ijЩ������Ҳ���յ�ALA�ϳ�ø����ͱ��ס��һ�ù�ص�ҩ����յ�ALA�ϳ�ø�ĺϳɡ�
2.ALA��ˮø����������ø��ALA��ˮø����������ø���ؽ������У���Ǧ�ж���������Щø��ʹѪ���غϳɼ��١�
3.��Ѫ�������ӣ�Ŀǰ�ѷ��ֶ�����Ѫ�������ӣ����ϵ(multi)һ����̼����ӣ�������ϸ��������ϸ������̼����ӣ�GM-CSF)����ϸ������3(IL��3)�����ٺ�ϸ�������صȡ����дٺ�ϸ��������(erythropoiefin,EPO)�ں�ϸ���������ֻ��з��ӹؼ����á���EPO����λ��7��Ⱦɫ�峤��21������4���ں��Ӻ�5����������ɡ�������Ķ�����193��������л���ɡ��ڷ��ڹ����о�ˮ��ȥ���ź��ģ���Ϊ166��������ij����ġ�������Ϊ18398��EPOΪһ���ǵ��ף��ɶ��ĺ��ǻ���������ɣ��ܷ�����Ϊ34000���ǻ���Epo�ϳɺ���ڼ�������Է��������Ҫ���á�����Ѫ��Epo��Ҫ������ϳɣ�̥������������Ҫ�ɸ���ϳɡ���ѭ��ѪҺ�к�ϸ���ݻ����ͻ����ȱ��ʱ��������Epo���ӡ�Epo�ɴٽ�ԭʼ��ϸ������ֳ�ͷֻ��������к˺�ϸ���ij��죬���ٽ�ALA�ϳ�ø���ɣ��Ӷ��ٽ�Ѫ���ص����ɡ�
��������Ѫ���صĺϳ��дٽ����á���Ѫ�����ֶ��鵰�ĺϳ��дٽ����á�
Ѫ���غϳɴ�л�쳣������߲�����������ǰ����Ķѻ�����Ϊ߲��֢(porphyria)�������Ժ�ϸ��������߲��֢(congenitalerythropoietic porphyria)������������ȱ����߲��ԭ��ͬ��ø����ʹ��״����������߲��ԭ���ת�����裬��ʹ��߲��ԭ���������ࡣ���������д�����߲����ͷ�߲������֡�
���������ϸ���Ĵ�л�ص�
�����ϸ��������ϸ���ˣ�����Ҳ�������塢�˵������ϸ���������ܽ��к���͵����ʵ�����ϳɣ�Ҳ���ܽ���������������������֬���ᡣѪ������Ψһ����Դ����ϸ����ȡ��������������ɢ���������ȵ��ء������ϸ�������Ĵ�лͨ·��Ҫ�������ǵĽͽ����������ͨ·�Լ�2.3һ�����������֧·(2��3-biphosphoglycerate��2.3�BPG)��ͨ����Щ��л�ṩ�����ͻ�ԭ��(NADH��NADPH)�Լ�һЩ��Ҫ�Ĵ�л��(2��3�BPG)����ά�ֳ����ϸ����ѭ����Լ120���������̼������������ܾ�����Ҫ���á�
(һ)�ǽͽ�
ѭ��ѪҺ�еĺ�ϸ��ÿ������Լ30g�����ǣ�����90��95%���ǽͽⱻ���á�һ���������Ǿ��ͽ�ɲ���2����ATP����ϸ�������ɵ�ATP��Ҫ����ά�ֺ�ϸ��Ĥ�ϵ����ӱ�(�Ʊá��Ʊ�)���Ա��ֺ�ϸ��������ƽ�⣻ά��ϸ��Ĥ�����ԣ������ĺϳɼ�������IJ��Ⱥϳɵȡ�ȱ��ATP���ϸ��Ĥ��������ƽ��ʧ������ϸ����Na���������K���ų���Ca�����������࣬��ϸ�����������ˮ�ֶ�������״�������ѡ�ͬʱ����ATPȱ������ʹ��ϸ��Ĥ�������½���Ӳ�����ߣ��ױ�Ƣ���ƻ��������Ѫ��
��ϸ�������ͽ������ɵ�NADH��H���Ǹ���Ѫ�쵰��ԭø�ĸ������ӣ���ø������Ѫ�쵰��ԭΪ���������ܵ�Ѫ�쵰�ס�
(��)2��3-�����������(2��3-BPG)֧·
���������ͽ�ͨ·�У�1��3-�����������(1��3-BPG)��15��50%�ڶ�����������λø��������2��3-BPG�������پ�2��3-BPG����ø������3����������ᡣ����2��3-BPG�IJ�֧ѭ����2��3-BPG֧·(ͼ10��19)��

ͼ10��19��2��3-BPG֧·
��ϸ����2��3-BPG����ø����Զ����BPG��λø��ʹ2��3-BPG�����ɴ��ڷֽ⣬�����ϸ����2��3-BPG��Ũ�ȴ����л����������۷壬���ǽͽ������м������л��������߳������������ٱ�(��10��4)��
��10-4����ϸ���и����ǽͽ��м�����Ũ��(�˷���/����ϸ��)
| �ǽͽ��м���� | ����Ѫ | ����Ѫ |
| 6-���������� | 30.0 | 24.8 |
| 6-������� | 9.3 | 3.3 |
| 1��6��������� | 0.8 | 1.3 |
| ������� | 4.5 | 5.0 |
| 3-��������� | 19.2 | 16.5 |
| 2-��������� | 5.0 | 1.9 |
| ����ϩ��ʽ��ͪ�� | 10.8 | 6.6 |
| ��ͪ�� | 87.5 | 143.2 |
| 2��3-DPG | 3400 | 4940 |
2��3-BPG���������ȥ��Ѫ�쵰��(deoxy Hb)��ϣ�2��3-BPG����Ѫ�쵰�צ�2��2�ľ������Ŀ�϶�������ǻ�֮�䣬�������������5����������������ǻ��Ĵ�����������л����μ��������ϣ�ʹ�������ǻ����ַֿ���״̬������ʹѪ�쵰���ɽ���̬���ɳ�̬ת�����Ӷ�����Ѫ�쵰����������(ͼ10��20)��
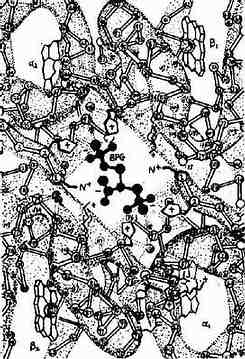
ͼ10��20��BPG��Hb������ʾ��ͼ
����ϸ����2��3-BPGŨ������ʱ������HbO2��������2��3-BPGŨ���½���������Hb������ϡ�BPG��λø��2��3-BPG����ø��pHֵ���ڡ��ڷ���ëϸѪ��ѪҺpH�ߣ�BPG��λø�����ƶ�2��3-BPG����ø����ǿ��ʹ��ϸ����2��3-BPG��Ũ�Ƚ��ͣ�������Hb��O2��ϡ�
��֮����������֯ëϸѪ���У�ѪҺpH�½���2��3-BPG��Ũ�����ߣ�������HbO2��������˵���������������ã�������Ҫ�������塣��2��3-BPG���������Լ���һ��ATP������Ϊ���۵ġ�
(��)��������ͨ·
��ϸ�������������ǵ�5��10%ͨ����������ͨ·��л��Ϊ��ϸ���ṩ��һ�ֻ�ԭ��(NADPH)��NADPH�ں�ϸ��������ԭϵͳ�з�����Ҫ���ã����б���Ĥ���ס�Ѫ�쵰��ø�����ϻ�������������ԭ����Ѫ�쵰�ȶ��ֹ��ܡ�
1.GSH����Ҫ���������ǶԿ����������ϻ���������ϸ���ڿ��Է�������������������(O��2)��ͬʱ��Ⱦʱ�İ�ϸ������������ɲ���O��2���ɱ����������绯ø(superoxidedismufase SOD)�������ɹ�������(H2O2)��

��GSH�ڹ����Ĺ�����ø�����½�H2O2��ԭΪH2O��GSH����������Ϊ����������(GSSG)�������ڹ����Ļ�ԭø���£���NADPH��H���������»�ԭΪGSH��(ͼ10��21)��
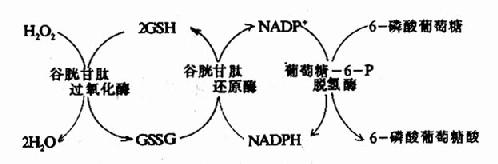
ͼ10��21�������ĵ������뻹ԭ
��NADPH���ɵĹؼ�øΪ������-6-��������ø����øȱ�ݵIJ���һ���������֢״�������������(���ʳij�ֲ϶�)Ӱ�죬��������Ѫ����Բ϶����յ������������ֲ��ֳƲ϶�����
2.����Ѫ�쵰��(methemoglobin MHb)�Ļ�ԭ�����ڸ����������ã���ϸ���ھ���������MHb�����������ں�ϸ������һϵ��ø�ټ���ø�ٵ�MHb��ԭϵͳ(��10?)����������ϸ����MHbֻռ1-2%��
��10��5����ϸ����MHb��ԭϵͳ
| ��ԭϵͳ | ռ�ܻ�ԭ�����İٷֱȣ�%�� |
| ø�ٻ�ԭϵͳ | |
| NADH����ø�� | 61 |
| NADH����ø�� | 5 |
| NADH����ø | 6 |
| ��ø�ٻ�ԭϵͳ | |
| ����Ѫ�� | 16 |
| ������ | 12 |
�ɱ�10-5��֪����MHb��ԭ����Ҫ��NADH-����ø����øΪNADH��H����NADPH����ø��(��NADPH��H��Ϊ��ø)Ҳ����MHb�����������ý�С������֮�⣬����Ѫ���GSH��ֱ�ӻ�ԭMHb���������Ϳ���Ѫ���GSSG�Ļ�ԭ����������NADPH��H�����⡣