�ڶ��ڡ�������ڷ��ӵĽṹ����
����ϵͳ�ļ���Ϊһ�ּ�����Ӧ�����ܵ����ֵ��ڷ��ӵ��ϸ���ƣ��䷴Ӧ�ij̶Ⱥ͵�һ�ɷֵķ�Ӧ���������ﷴ���������¶����еģ��Ӷ������˻��������ά�ֲ���ˮƽ��ƽ�⡣�������ð����������棬�������Ա�ʧ�һЩ�������������á�ǰ��ָ�ѻ�IJ�����Ӿ����ȶ����粻��ʱ���ϸ��Ĥ��ϼ�Ѹ��˥��ʧ�������ͨ������������ö�ʹ�ѻ�ķ���ʧȥ���ԡ���һ���н��漰��һ�����档
һ��C1������
C1�����C1INH����Ѫ���и߶��ǻ�����һ�ֵ����ʣ��������ߴ�35-49%�������Ranoff��lepow(1957)�����֣�����ΪC1��ø���Ƽ�������ͬʱSchultze�������Ϊ��2�����ǵ��ס�C1INHΪһ�������ӣ���478��������л���ɣ�������Ϊ104kDa����478��������л���ɣ�������Ϊ140kDa�����������Զ������ͼ5-13����C1INH���ڵ���Ҫ��ʽ�ǣ�����C1r��C1s����γ��ȶ��ĸ����������C1˿���ᵰ��øʧ������û����ǣ�C1INHͨ���ṩһ������C1r��C1s���������������Ϊ���������bait��������c 1r��C1s�ѽⱩ¶��һ�����Բ�λ��Ȼ������C1r��C1s����γɹ��۵������������������á����⣬C1INH���ɷ�ֹ��ȱ������ʱ��C1�Ժܵ͵�����һ�����ʳ��ֵ��Է������������£�ѪҺ�еĴ����C1�ɱ�7������˷���Ũ�ȵ�C1INH����ϣ��Է�ֹC1���ڹ���ı��������Է������C1ͬ��ԭ�����帴����Ľ�ϣ���ʹC1��C1INH�����������ж����͡������������⣬C1INH����������Ѫ���Ӣ�a����a�������ͷ�ø������ø�����������Ѫ�����ĺ�����ϵͳ��Ҳ����Ҫ�ĵ������á�
����C1INH cDNA���еķ������֣�C1INH����������˿���ᵰ��ø�����serpin���������Ա����1���ȵ���ø����1���ӵ���ø������Ѫø��ȣ�Լ��30%�İ�����ͬԴ�ԣ��������C�˵�120�������ᡣC1INH�ı������λ�ڵ�11��Ⱦɫ��Ķ̱�11.2������13������C1INH������ȱ��ʱ���ɵ����Ŵ���Ѫ������ˮ�ף�hereditary angioneurotic edema,HAE�������걨��Ӧ��C1INHŨ��������HAEЧ�����ã�����δ�㷺�����ٴ���
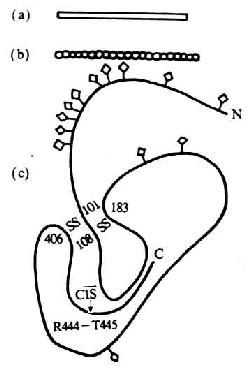
ͼ5-13��C1 INH���ӵĽṹģʽͼ
ע����a��Ϊ�羵�۲�ģʽͼ
��b��Ϊ����ɢ��ģʽͼ
��c��Ϊ��ɫ�����Ķ����ṹģʽͼ
����C4��ϵ���
C4��ϵ��ף�C4bp����һ�ֺ����ḻ�Ŀ�����Ѫ���ǵ��ף�������Ϊ550kDa��1977����Ferreira��������������ӽṹģʽ�ֶ���Dahlback�ȣ�1983�������ġ�֩��������spiderlike���ṹ��������ṹ�����ܡ�C4bp��8���ǵ�λ��ɣ��羵�¹۲�����֩�룬������7����������ͬ����Ϊ70kDa���ij���������������549��������л���ɣ������Զ���������ᣬ����ͬ������л���ɣ������Զ���������ᣬ����ͬ������һ�����塣��������8��SCR����N������ͷ����Ϊ��C4b���ϵIJ�λ������λ�ڵ�332-395λ��������л�����8�������������϶̣�45kDa����235��������л���ɣ�����4��SCRΪ�뵰��S��PS�����ϵIJ�λ��ͼ5-14����C4bp�����ַ�ʽ���Ʋ���Ļ����һ�ַ�ʽ�ǣ�ͨ������C2����C4b������c 4b2a��ȡ��C2a����ͨ������C4b�Ľ�϶���ֹʣ���C2ͬC4b��ϣ��ɴ�����C3ת��ø���γɡ�C4bp��C4b�Ľ��������ϸ������C4b���ӵ��������ȣ��ҽ�C2ͬC4b�Ľ��������27�����ڶ��ַ�ʽ�ǣ�C4bp��ΪI���ӵ�һ�ָ������ӣ��ٽ�I���Ӷ�C4b���ѽ⡣��C4bp����ʱ��I���ӿɽ�C4b��a`����ȫ�ѽ⣻��C4pbʱ��I���ӵ��ѽ����ò���ȫ������I���ӽ�ϵĻ��Բ�λλ�ڵ�177-322λ������л�����PS��C4pb�ĵ�8��������������ϣ���Ӱ��C4bpͬC4b�Ľ�ϣ������ӳ�C4bp�İ����ڣ��Ӷ�ǿ��C4bp���������á�

ͼ15-14��C4bp�Ľṹ��ģʽͼ��
C4bp����λ���˵ĵ�1��Ⱦɫ�峤��32����ͬCR1��CR2��H���ӡ�MCP��DAF�ȵĻ���������������������Ŀ��ͬ��SCR��
������˥�����ӣ�CD55��
��˥�����ӣ�decayaccelerating factor,DAF����Nicholson-Weller�ȣ�1981������������ȡ�����Բ��������˺������ϸ�������д�����һ��Ĥ���ס�������дٽ�C3ת��ø�Ա�Ļ��Թ��������ڻ�ԭ��������SDS-PAGE���Թ�����-Schiff�Լ�Ⱦɫ������������DAFΪ����Ĥ�ǵ��ס��˺������DAF��������ͬ�ֱ�Ϊ70kDa��60kDa�����Ѱ���ϸ���ֻ���ԭ�����ΪCD55��Davitz�ȣ�1986��ͨ������֬��������PI�������Ե���֬øC��PI-PLC������������Ѫϸ�����ͷ�DAF����ʵ̽����DAF�Ǿ�����֬��������glycosylphodphatidylinositol,GPI��ê���̶���ϸ��Ĥ�еġ����ǵ���Cĩ�˹��۽���ں�PI������֬�ϣ��پ�PI����ϸ��Ĥ֬��˫������СҶ�С��о�������ĤDAF��Ǩ���ʽӽ���Ĥ��֬��Ǩ���ʣ��ȴ����Ĥ����Ǩ���ʸ�һ������������Ϊ�������ڴٽ���Ŀ����ĤDAF������ϸ�����������C3b��C4bƬ�νӴ�������DAF������֬�������ṹ�����ܾ���ת��ϸ���źŵ����á�
��Nicholson-Weller��֤ʵ�ķ�����Ϊ70kDa��ĤDAF�⣬Kinoshita�ȣ�1987����Western blotting���˺�ϸ�����滹���������Ϊ140kDa��һ��ĤDAF������ΪDAF-2��DAF-2��Ĥ�ϵ���Ŀ����70kDaĤDAF��1/10����Ҳ�дٽ�C3bת��ø˥��Ļ��ԣ�Ҳ����GPIê�ṹ������DAF-2�ķ�������70kDa��ĤDAF��һ������ʾ��ΪĤDAF�Ķ����壬���ö��ϻ��Ҵ�����SDSʹ�����ԣ������ܽ�DAF-2�ѽ�������ɷ֣���DAF-2�ľ�ȷ�ṹ���д���һ�����������⣬����Ӧ����λ��RIA�ⶨ������Ѫ������Һ����Һ����Һ����ĤҺ���Լ�Һ���Լ���֯���������о���������Ե�DAF��sDAF����ˮƽ��40-400ng/ml��Χ����������Һ�е�sDAF�������Ե��ں�ϸ����ĤDAF�ķ���������ˮ��Ҳ��ĤDAFС��������ϸ������C3ת��ø����װ��Ļ��Խ�ĤDAFԼ��100�������Ծ��дٽ����γɵ�C3ת��ø˥������ã�Ч��������C4bp��
ĤDAF�㷺�ֲ��ڸ���Ѫϸ��������������ϸ���ϣ�������ϸ������ϸ��������ϸ�����ܰ�ϸ����T��B����ѪС�塢���赥����ϸ������ϸ����ʼ��ϸ������Ĥ�����Ĥ��������ճĤ��������١���С�ܡ����ס��ӹ�ճĤ����Ĥ���İ�����Ĥ����Ƥϸ�������ӣ��Լ��������꾲����Ƥϸ���ϡ���NKϸ������ȱ�硣����Ѫ�쵰����paroxysmal nocturnalhemohlobulinuria,PNH�����˵ĺ�ϸ����Ҳȱ��DAF������ȱ���̶Ƚ��ò���Ϊ�����͡�PNH���˵ĺ�ϸ���Բ���鵼����Ѫ���ø߶����У��������ں�ϸ����ȱ��DAF��������GPIê�ķ��Ӷ�����ġ�DAF����Cromer��ԭ����������֮ΪInaba��IFCȱ����Cromer��ؿ�ԭ�ĸ���Ҳȱ��DAF��
DAF����ѧ���Լ�����������ʭ�����֤ʵ�����ɱ�������ϸ�����ⲹ��鵼���ܽ��ƻ��������û����ǣ�DAF��������ֹ��������;��C3��C5ת��ø��װ�䣬���ҿ�ͨ���յ�����λC2a��Bb�Ŀ��ٽ����ʹ���γɵ�C4��C5ת��øʧȥ�ȶ��ԣ��Ӷ����Ʋ��幥����λ�Ļ��ͼ5-15����DAF�������������ý�����ֱ�ӽ����ϸ���ϵ�C3��C5ת��ø��Ҳ��DAF�����ư�ϸ���������IJ��弤�������������߸������DAF������ΪI�����ѽ�C3b��C4b�ĸ����Ӷ��������á����⣬DAF�䲻����ֹC2��B���ӣ��ֱ�ͨ����C4b��C3b��ϣ���ϸ���������ϣ���ȴ��ʹC2a��Bb�����ǽ�ϵIJ�λ����������Զ���ֹC3ת��ø��װ�䡣
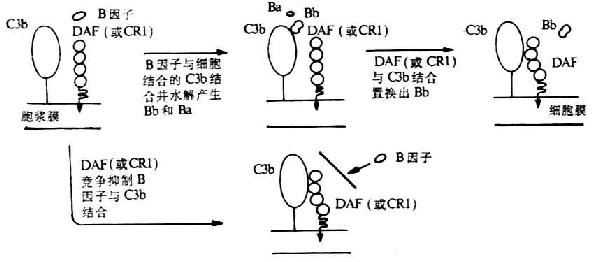
ͼ5-15��DAF�������;����C3ת��ø�γɵĵĻ���
������DAF�Ļ���λ�ڵ�1��Ⱦɫ��ij�����32��һ��800kbƬ���ڣ����������ֲ��弤����ڼ���RCA���Ļ����������������˳������Ϊ��MCP-CR1-CR2-DAF-C4bp������DAF����ij���ԼΪC4bp������DAF����ij���ԼΪ35kb����������ø��������Ϊһ������������DAF����ķDZ���������3��������ø��Ƭ�γ��ȶ�̬�ԣ�RFLP���ṹ������ΪHind��ø��λ�㣬1��ΪBamh ��ø��λ�㡣DAF��cDNA�ѿ�¡�ɹ����������˺�����Ͱ��������з���������DAf cDNA��Ĥ�ǵ��Ľṹ��ͼ5-16��DAF��cDNA�ṹ��5`�˿�ʼ����Ϊ���ź��������ĸ�SCR������1143bp��������˿�������հ��ᣨS/T��������Լ70�������ᣩ����ˮ���������ֹ��3`�˵�poly��A������cDNA�Ƶ��İ��������еó�DAF������381������������ɣ�����34����������ź��ģ�����S/T����ΪDAF�д���������O-���ӵ��ǻ�����λ��SCR����1��N-���ӵĵ��ǻ�����λ��Cĩ�˵���ˮ���ڷ��������֬��ȡ����ΪDAF������ϸ��Ĥ���ϵIJ�λ��
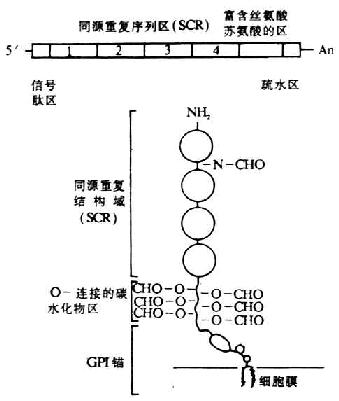
ͼ5-16��DAF cDNA����Ĥ�ǵ��Ľṹ
�ġ�Ĥ�����ף�CD46��
Ĥ�����ף�membranecofactor protein,MCP����Cole�ȣ�1985��Ӧ��C3b�Ͳ���������Ѫ�ܰ�ϸ���Ϸ��ֵ�һ��Ĥ���ס����������صĵ�Ӿ���������������Ϊgp45-70������һ���о����֣�����I���ӽ鵼�Ķ�C3b��C4b���ѽ��и������ԣ��ʸ���ΪMCP������MCP�Ķ��¡��������Ա����ӣ�DAF����H���ӻ�CR1������Ӧ���Ӷ���Ϊ���Dz���ϵͳ��һ���µĵ��ڵ��ף��ڵ��Ľ���ʰ�ϸ���������ۻ��Ͻ�������ΪCD46��
MCPΪһ������Ĥ�ǵ��ף�������45-70kDa������RCA����صij�Ա��Ҳͨ��GPIê�̶���ϸ���ϡ�MCP��ϸ���ֲ����㣬������ϸ����ѪС�塢Tϸ����Th��Ts��Tc����Bϸ����NKϸ������Ѫϸ��ϵ������άϸ������Ƥϸ������Ƥϸ������״����ϸ���ȡ�����ͬ���͵�ϸ���ϱ��������������ͬ������Ѫ������ϸ������ϸ��Ϊ1���/ϸ������Ѫϸ��ϵΪ2-5���/ϸ����Hela��Hep-2ϸ���ֱ�Ϊ10���/ϸ����25���/ϸ�������ֱ��������IJ��죬���ܷ�ӳ��MCP������ϸ��������ϸ���ϲ�ͬ�ķֻ��ͻ״̬�����⣬���ڲ�ͬϸ���ϱ����MCP��ͬ����˿ɵ�����������;����C3ת��ø�γɡ���һ��������Ҫ����ΪC3������ת��tickover���Ļ��ƣ��ܹ��������ϵIJ���C3b��C4b���п����γ�Խ��Խ���C3ת��ø�������ڴ��������ϸ�����и�ˮƽ��MCP����˿ɱ�������ϸ�����ⲹ��鵼�����ˡ��෴����������������²�������ȱ��MCP���������������DZ����C3b��ɵ��Ա���������C3b��B���ӽ�ϵ�����������H���ӽ�ϵ��������Ӷ��ٽ�c 3bBb��������γɣ���������Щ��������ϲ�����Ч�ػ�����ս����ƻ���������⣬ϸ��������Һ��ĺ���������H���Ӻ�B���ӵ������йأ���Һ�Ậ���ϸߵ�ϸ����H���ӵĽ������������B���ӵĽ����������Һ�Ậ���ϵ͵�ϸ��������෴������ϸ�������Һ��ĺ������ڲ����ද����˵�ϸ�����������������ͬB���ӽ�϶����²������;���ļ��

ͼ5-17��MCP����I�����ѽ�C3b�Ļ���
MCP��Ϊһ�ֲ�����ڵ�������Ҫ������ѧ���ܡ��ڵ�����ǿ���£�������C3b��C3bi��ϣ�����CR1�������͡�MCP����Ҫ�������丨���ӻ��ԡ�������C3b��C4b��϶��ٽ�I���Ӷ�C3b��C4b���ѽ���ͼ5-17�����Ӷ�������������ϸ�����ⲹ��鵼���ܽ��ƻ������˽�MCP���������ó�Ϊ��Դ�Ը����ӻ��ԡ�Seya�Ȼ�����MCP������ǿC3ת��ø�Ļ��ԣ���������;����C3ת��ø���������������в����ˡ�CR1��H����Ҳ�Ǿ��и����ӻ��ԵIJ�����ڵ��ף���H���ӵ���һ���Խ�ΪMCP��1/50��
��MCP�Ļ���λ�ڵ�1��Ⱦɫ�峤��32������������Ϊ43kb������14�������Ӻ�13���ں��ӡ�Seya���ô�HSB-2Tϸ��������õ�MCP����ĩ�������һ��17mer�ķ���Ѻ�����̽�룬�Դ˴�U937��cDNA�Ŀ���ɸѡ��һ����1.5kb��cDNA���������������cDNA�к���һ��43bp��5`�˷DZ�������һ������384��������Ŀ���������ǰ34��������Ϊ�ź��ģ���350��������ΪMCP�Ķ�������������Ϊ39kDa���ڶ�������ǰ250���������У�����4�����ڵ�CSR��SCR��Ϊһ�θ���˿���ᡢ�հ��������л���29��������Ƭ�Σ������Ǹ߶�O-���ӵ� �ǻ���λ�㣩���ٺ�����Ϊ13��������Ĺ��ܲ�������24��������Ŀ�Ĥ����105��������İ���ê��23�������ṹ�ɵİ���β���������MCCP�ı�������ڸ���˿����/�հ��ᣨS/T�����Ͱ���β�����ں�����������MCP��DAF����ͬԴ�ԡ�
�塢H����
H������Nilson�ȣ�1965�����֣����ݵ�Ӿλ��������Ϊ��1H����Whaley��Ruddy��������ΪC3b�����������ӡ�����ȷ����Ϊ��1213����������ɵĵ����ǵ��ף�������155kDa�����г���״���֣�Ҳ�����������½�����羵��鷢�֣�H���ӵ�Ӱ��Ϊһ������˳�ķ��ӡ��쳤�ͷ��ӳ�49.5nn�����ֱ��Ϊ3.4mm��������������Ͷ����۵�״����˷��ӵ�ʵ�ʳ��Ƚ�Ϊ��չ�͵�һ�룬���乹����ʶ�������ͨ����ɫ����������H���Ӽ��ަ�����Ҳ�ަ��۵���������ά���书�ܻ��ԵĹ���H���ӵĹ��ܰ������¼������棺��1��Ϊ�����ӵĸ������ӣ�������C4b�Ԣ����ӵ������ԡ�����H����ʱ����������C3b�Ľ�ϳ�˿״��������H���Ӵ���ʱ��I������C3b�Ľ�ϱ�Ϊ��˿״��ͬʱ��C3b�Ľ��������ǿ������H����ʱ���ٸ�15������H����ǿ��I���ӵĻ�����������H������C3b��Ϻ�ʹC3b����ijЩ����仯����������I���ӽ�ϵ�������H������C3b��ϵĻ��Բ�λ��������N��35kDa���֡���2������C3ת��ø���Ա䣺H�����ܽ���ͬC3b��ϵ�B���ӻ�Bb��C3ø���������ʹ֮ʧȥø���ԡ���3����ֹ���;���г�ʼ�ͷŴ�C3ת��ø���γɡ���֤ʵH���Ӻ�B������C3b����ͬһ��ϲ�λ����H���ӿ�ͬB���ӻ�Bb������C3b�Ľ�ϡ�����H���Ӵ���ʱ��B���Ӳ�����C3��H2C����C3b��ϣ���˲����γ�C3��H2C��Bb��C 3bBb����H���ӶԹ����Ϻ�Һ���е�C3b�����ڲ�𡣶�Һ���л����ڷǼ�����������ѽ⡣�����ڹ̶�������������ĸ���ǵȣ������C3b��H�������C3b��������B�����൱�����߾����Ľ�������γɲ���C3ת��ø���Ա�֤���;���Ļ�����о���������ϸ��Ĥ������ǿC3b��H���������Ļ�ѧ�ɷ���������ذ������ǡ����ڴ����ϸ������ȱ�����ᣬ�����Щϸ���������ɻ���;���������������ڶԸ�Ⱦ�Ŀ��ơ���4��������P���ӻ��������ӣ�NeF������γ��ȶ���c 3bBbP��C 3bBbNeF��H���Ӷ�����Ҳ��һ�������ã�����Ч��Ҫ��CR1��öࡣH���Ӷ�C5ת��ø��c 3bnBb��C 3bBb3b���Ļ���Ҳ���������ã�������C5�������C3bʹC5�����ѽ⡣���⣬���귢��H���ӻ����յ�����ϸ������IL-1��������Ӧ��ĵ��ڡ�
����H���ӵĻ���λ���˵ĵ�1��Ⱦɫ��ij���32�������ж�̬�ԡ��ѷ�������5�������ͣ���FH1-5��H���ӵĺ����������ѽ����˼��������Ƶ�����ȫ���������һ���ṹ������20��SCR�����C3b��ϡ�H������MCP��CR1��CR2��DAF��C4bp����ͬԴ�ԣ���ͬ���ڲ��弤����ڼ���RCA���������ij�Ա��
����I����
I���ӣ��ɳ�C3bINA��Ϊ��Դ������Ѫ�嵰�ף���˫��״�ṹ������ȫ��13nm������С��L����4.9nm������˿���ᵰ��ø���ԣ�����H����5.4nm����C3b��ϡ�I���ӵķ�����Ϊ88kDa������50kDa������38kDa�������Զ���������ӡ�I���ӵ�����ѧ�����ǣ���C4bp��MCP��H���Ӻ�CR1�ȸ������ӵ�Эͬ�£���C4b�ѽ�ΪC4d��C4c;ʹC3b�ѽ��C3f�γ�C3bi,�����ٽ�һ���ѽ�ΪC3dg��C3c���ɴ˶����Ʋ���ϵͳ�Ļ��
������I���ӵĻ���λ�ڵ�4��Ⱦɫ���ϡ�����I���ӵ�cDNA���з������֣�������Ϊ˿������ø�Ļ����������ӵ���ø���ȵ���ø�����Ե���ø����ͬԴ�ԡ����������Ǿ��и������װ���Ĺ���������C8��C9�͵��ܶ�֬��������Ⱦ���ͬԴ�ԡ�I���ӽṹ�����ͻ�䣬�ɵ���������I����ȱ�ݣ����C3�Ĺ���������������Ⱦ��Ѫ����ˮ�ס�
�ߡ�������������
��������������AI���ֳ�Ѫ������øN��������310kDa����8����ͬ�Ķ�������ɣ�ÿ����������36kDa��AI�����Ȼ���ø�Ļ��ԣ���ȥ��C4a��C3a����C5a Cĩ�˵ľ�����л���ʹ��ЩƬ��ɥʧ��������ػ��ԡ�
�ˡ�S����
S���ף�CP��S��ΪѪ����һ�֦������ǵ��ף�������83kDa��SP����Ҫ���������ǿ���C5b��7������̬��ϲ�λ������ϸ��Ĥ֬�ʣ�ͨ���γ���ˮ�Ե�SPC5b��7����дΪS5b��7���������ʹC5b��7ʧȥĤ��ϻ��ԡ���������ɱ���������λ�ڽ���ϸ������żȻ�Ĺ�����������ˮ�Ե�SC5b��7���ɼ�����1�����ӵ�C8��3�����ӵ�C9��ϣ��ֱ��γ�SC5b��8��C5b����9��3�������C9�ۺ��γɿ����Ӷ��ɱ���������λ�ڽ���ϸ���������ܲ���Ĺ��������ˡ�SP��C8��C9�Ľ�ϲ�λΪ�����ַ����и������װ���Ĺ��ܹ��������羵�¹۲죬SC5b����9��3�������һШ�νṹ��SPλ��Ш�εĿ������ڸDz��嵰����ˮ�����Ӷ����MAC��Ĥ��ϲ�λ�����⣬2��3�����ӵ�SP��C5b��7��C5b��8������Ľ�ϣ�����ʹ��Щ���������ܣ�������ˮ����ˮת����SPҲ������Ѫ���̣�ͨ�����ſ���Ѫø�����Ѫø�������������Ѫø��
������SP����λ�ڵ�17��Ⱦɫ��ij����ϣ���cDNA�ѿ�¡�ɹ����������з����������������ϸ��ճ�����õIJ���ճ�����ף�vitronectin����������ȫ��ͬ����֤��������ͬһ���ס�
�š�CD59
CD59�ǽ��꣨1989���ŷ��ֵ�һ�ַ�����ֻ��18��20kDa��Ĥ�Ե��ڵ��ס���Sugita�ȣ�1988�꣩�������������в�ͬ�����ƣ���ͬ����������20��homologous restriction factor-20��,�����أ�protectin����Ĥ��Ӧ�����������membraneinhibitor of reactive lysis,MIRL���ȡ�CD59��103��������л���ɣ����е�һ��N���ǻ���λ�㣬��C�˽�GPIê�̶���ϸ�����档CD59�ֲ����㷺����֤��Ƥ�����Ρ������ȡ��Ρ����١���ϵͳ��̥���Լ�����Ѫϸ������ϸ�����ܰ�ϸ����������ϸ����ѪС�壩�;����Ͼ��б��C59����Ҫ���������ǣ���ֹMAC��ͬ�ֻ�����ϸ�����ܽ��ƻ�����ͬ���������ã�homologous species restriction,HSR)�����û���Ϊ��ͨ������C7��C8��C9�Ľ�϶���ֹMAC��װ��������C5b��7�������Ϻ���ֹ������C8��ϣ�������C8��Ϻ�����谭�����MAC�ϵĵ�1��C9���ӵ���չ���Ӷ�����ֹ���C9�Ľ�ϣ���������MAC�е�C9��Ϻ��ֿ���ֹC9��һ�����ӵľۺϡ�����������CD59���ڵ�����£���ͬ�ֻ�����ϸ���ı���㲻��˳�������MAC��ȫ����װ��������װ��C5b����9��n������Ӷ������˶�ͬ�ֻ�����ϸ�����ܽ����á�Rooney�ȣ�1991���ı���������CD59������ͬ���������õĻ����Ա����ӹ��ڵ�̥������MAC�Ĺ���Ҳ�����á����⣬CD59���ٽ�Tϸ��ճ���뼤�ά�־�����Ů����ֳ���ڵ��ȶ�������ԣ��Լ�����PNH�������������á�
CD59��cDNA�Ѿ���¡����ȷ��CD59�Ļ���λ���˵ĵ�11��Ⱦɫ��̱ۣ�������������백�����������Ӧ������С���CD59��ͬԴ�ԡ�
ʮ��SP40/40
��1982���������ѧ�����������Ⱥ����һ������������Ƕ��ǵ����ʣ��ҽṹ��ͬ������������벹���ĩ�˳ɷ���ء���Щ�ɷ��в�ͬ�����ƣ���Clusterin��SGP-2��SP40/40��TRRM-2��T64��GP111��SP80��CLI��apo-J��NA1/NA2�ȡ�Fritz�Ƚ�����Clusterin������ΪȺ���أ���������
Ⱥ���صĵ�����������SP40/40��һͬԴ���з��ֵġ�SP40/40����70���ĩ�������˾����з��ֵģ��̺�����С�������IC��ѪҺ��MAC��Ҳ����������ڻ�������ĩ�˲��帴��������ڣ�����Ϊ����ĩ�˲��帴����ij�Ա֮һ��SP40/40ΪѪ���еĦ��ǵ��ף�������80kDa���ɷ�������Ϊ40kDa�Ħ��ͦ��ǵ�λ����ɣ����Գ���ΪSP40/40����֪SP40/40��427��������л�����ɣ����ǵ�λΪ222��������л������ǵ�λΪ205��������л������ͦ��ǵ�λ��3������������Ӷ�����һ������塣�����ǵ�λ�İ�������ɳ��ʰ����⣬����������ƣ����������������������ԡ���Ѫ����SP40/40�ĺ���Ϊ35��105��g/ml��ƽ��62��g/ml���������˾�����ȴ�ɸߴ�15mg/ml����Ϊ���뱣�����ӵĻ����йء�
SP40/40��MAC��װ�ĵ��ڵ��ס�������C5b��7��C5b��8��C5b��9��ϣ���MAC����װ���������ã��Ӷ��ɷ�ֹMAC����ϸ�����á�����SP40/40��S��������Эͬ���ã���ʹMAC��Ϊ�����ԵĶ�ʧȥ��ϸ�����á���֪C8��SP40/40�Ľ�ϲ�λ��C8�¡�C9��SP40/40�Ľ�ϲ�λ��C9b��
ʮһ��ͬ����������
ͬ���������ӣ�homologousrestriction factor,HRF�����ֳ�C8��ϵ��ף�C8bp����ΪZalman�ȣ�1986����C9-Sepharose�Ͳ������˺�ϸ��Ĥ�Ϸ����һ������C8��C9��ϲ�����C9��ͬ�������Ե�Ĥ���ף�ͨ��GPIê�̶���ϸ��Ĥ���档���ʺ�ϸ��Ĥ�ϵ�HRF������Ϊ65kDa�������䶳�ij¾��Ժ�ϸ��Ĥ����Ϊ38kDa��HRF�����������˵ĺ�ϸ����������ϸ��������ϸ�����ܰ�ϸ����ѪС���ϡ�HRF�Է�Ӧ����Ѫ���õ��ϸ�����������ԡ��羭��HPF���崦�����˺�ϸ������C8��C9�ķ�Ӧ����Ѫ�������������ߣ�����8�ֶ�����Դ��C8��C9�ķ�Ӧ����Ѫ���������С��������������HRF������C9��C8�Ľ�ϼ�C9�ۺϣ��Ӷ���ֹMAC��������ϸ����Ĥ֬��˫�㼰ϸ���ܽ⡣HRF���ܻ����ܰ�ϸ��ɱ��C9RP���鵼���˴�����ܰͶ����ϸ����PNH���ߺ�ϸ����ADCC���á�
��������11�ֲ�����ڷ��ӵ����Լ�����ѧ���Լ���5-2
��5-2��������ڷ��ӵ����Լ�����ѧ����
| ���ڷ��� | ��������kDa�������������� | Ѫ��Ũ�ȣ���g/ml����ϸ���ֲ� | ������������õķ��� | ����ѧ���� |
| Ѫ�嵰�ף� | �� | �� | �� | �� |
| C1INH | 104 | 200 | C1r,C1s | ��C1r��C1s���۽��ʹC1ʧ� |
| �� | ��serpin�� | �� | �� | ��C1�����ֹ���Է�������Ƽ����ͷ�ø������ø����Ѫ���Ӣ�a����a |
| C4bp | 550��12��SCR�� | 250 | C4b | ����CP�е�C3ת��ø�γɣ�����CP�е�C3��C5ת��ø˥�䣻��ΪI�����ѽ�C4b�ĸ������� |
| H���� | 155��20��SCR�� | 480 | C3b | ��ΪI�����ѽ�C3b�ĸ������ӣ�����AP�е�C3ת��ø˥�� |
| I���� | 88 | 35 | C4b,C3b | �ڸ�������Эͬ�£��ѽ�C4b��C3b |
| AI | 31������øN�� | 35 | C3a,C4a��C5a | ˮ��Cĩ�˾�����л������������� |
| S���� | 83��Vitronectin�� | 505 | C 5b��7 | ��C 5b��7����γ�SC5b��7��ʹ֮ʧȥĤ��ϻ��� |
| SP40/40 | 80��������壩 | 50 | C5b��9 | ����MAC����װ���������ӻ��� |
| Ĥ�����ӣ� | �� | �� | �� | �� |
| MCP��CD46�� | 40��70��4��SCR��GPIê�� | ����ϸ�����Ѫϸ������Ƥϸ������Ƥϸ�� | C3b,C3b | ��ΪI�����ѽ�C3b��C4b�ĸ������� |
| DAF��CD55�� | 70��4��SCR��GPIê�� | �������Ѫϸ������Ƥϸ����ճĤ��Ƥϸ�� | C 4b2a,C 3bBb | ����C3תø�γɡ��ٽ�CP��AP�е�C3ת��ø˥�� |
| HRF��C8bp�� | 65��GPIê�� | ��ϸ����PMN������ϸ�����ܰ�ϸ����ѪС�� | C8��C9 | ����C9��C8��ϼ�C9�ۺϣ���ֹMAC��������ϸ��Ĥ |
| CD59��MIRL�� | 18��20��GPIê�� | ��ϸ����PMN���ܰ�ϸ����ѪС�� | C7��C8��C9 | ͨ����C7��C8��C9��϶���ֹMAC����װ����ֹMAC�ܽ�ͬ�ֻ�����ϸ�� |