��һ�ڡ�������гɷֵķ��ӽṹ������
����ϵͳ��������;���У��漰��14�����嵰�ף�C1-9����B��D��P���ӣ��IJ��롣�����������ڷ����Ŵ�ѧ�ͷ��ӿ�¡������Ӧ�ã��Ѳ������ಹ����ӵĽṹ�����ܡ�����ϳɼ��Ŵ��������Ӷ���ٽ������ǶԲ���ϵͳ������̻�������ʶ�ͶԸ���������ӹ��ܵ������˽⡣
һ��C1����
C1�Ǿ��伤��;���е���ʼ�ɷ֡�������1�����ӵ�C1q��2�����ӵ�C1r��2�����ӵ�Cls��Ca2+���Ӷ��ɵĴ���Ӹ����������ԼΪ750kDa������C1qΪ����ʶ�����õ��ǵ�λ��C1r��C1sΪ���д����õ��ǵ�λ��
��һ��C1q
C1qΪ���ֲ�������з��������410kDa���Ħ��ס�����ӽṹ��������ӣ���A��B��C���ֲ�ͬ���͵���������ɡ�����A��B��C����6������18����A��B��C���������ķ�����������ͬ���ֱ�Ϊ24��23��22kDa��������222-226��������л����ұ˴�ͬԴ��ÿ�������ɺ����װ���л���һ���̵�Nĩ��������ɣ�����Ϊһ��81��������Ľ�ԭ���У����ظ�����������Gly-X-Y,Y��ͨ��Ϊ�Ǹ������������л����������е����ಿ��Ϊ�ǽ�ԭ�Եġ�A��B���估����C�������һ������������ӡ�18��������ÿ������ͬ���������һ�������������ʹ���6�������Ľṹ��ÿ����������������˿״��ԭ���ɷ���ɡ���6�������ṹC�����ڰ��������е�����������γ�6������״������ͷ�����ʻ�����չ�����ڽ�N��ԼΪ1/2ȫ���������ṹ����״��ƽ�����У���Nĩ��ΪC1q��β��������ڵ羵�¹۲죬C1q���ӵ�ͼ�����һ��ʢ���������㻨��ͼ5-1��
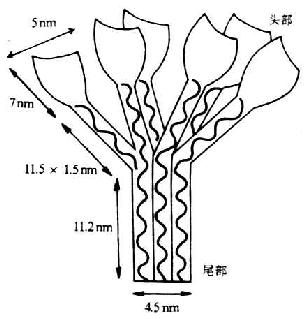
ͼ5-1��C1q�Ľṹ��ģʽͼ��
C1q�Ľ�ԭ�����н��C1r��C1s�IJ�λ����֤ʵ�ۺϵ�C1q�̼�Bϸ����ǿ�����Ig�����ã�Ҳ��ͨ����β������ɵġ�C1q�Ĺز�������ʶ��IgFcƬ���ϲ����ϲ�λ��λ�㣨C1q��C1q-R����ã���������6������ͷ���ʻ�����չ����������������Ig�Ӵ��Ļ��ᡣC1qͬ1�����ӵ�IgM��ϼ��ɱ������������ͬ����IgG���ӽ�ϲ��ܱ������������IgG������ϸ��Ĥ�ϵľ��벻������700nm��C1q����4��IgG����Ľ����������Ϊ��IgG3>IgG1>IgG2>IgG4��
Reid���Ѷ�C1q���ӵ�A��B�����˲��ְ�����������������A��B����cDNA��¡�����з�������ˣ�C1q���ӵĴ�һ���ṹ�Ѿ���ȷ������C1qA��B��C���������Ļ������λ���˵ĵ�1��Ⱦɫ��Ķ̱�34.1-36.1����
������Clr��Cls
Clr��Cls��Ϊ��һ���������ӣ��ֶ���˿���ᵰ��ø��ԭ����Clr��Cls ���������ɽӽ�700������������ɡ�λ��Cĩ�˵�Լ250��������Ϊ˿���ᵰ��ø�������ȵ���ø���ӵ���øͬԴ��ͬ��������嵰��һ�������Ƕ�����Ƕ��mosaic�����ף����ɲ�ͬ��������ɵĹ̶�������϶��ɣ����Һܿ��ܴ����������۵���������ṹ������module����
�羵�¹۲������Clr��Cls�ķ��ӹ��ͼ�Ϊ���ƣ�����һ�˴�һ����С������״���ӡ�
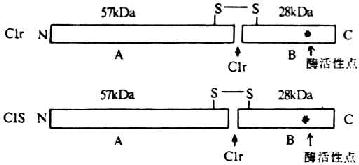
ͼ5-2��Clr/Cls���ӵĽṹ
�������ӵ�Clr��ͬ�ȷ��˵�Cls��Ca2+���ӳ�Ť���ġ�8�����Σ��̼���C1q��ͷ����6�������ṹ�䣨ͼ5-3����Clr��Cls�ķ������������ṹ�䣨ͼ5-3����Clr��Cls�ķ�������Ϊ85kDa�����Ǽ�����ڷ����ڵľ�������������л�����ѣ��γɷ������ֱ�Ϊ57kDa��28kDa��A��B����Ƭ�Σ����������Զ���������ӣ����������Ӳ�ĩ���롣��BƬ���Ϻ���˿���ᵰ��ø���Ե㣬Ϊ���Ӣ������ͼ5-2����AƬ������Clr��Cls���Ӧ�ĵĹ���������Ӧ�������������ģ���������λ����ࡣ��һ��C1INH��C1r����ţ���һ�������߸������ϵ�Clqʱ��C1INH���������ü����Ƴ�����ͨ��C1q�Ľ�ԭ�Ա�����ͷ�����ƶ����ݵ�������������Ӵ˴��ٴ��ݵ����������ӵ�C1r���յ�C1r���Ӱ��ı䲢�ѽ������C1r��C1r������������C1sʹ֮��Ϊ���C1s��C1s����
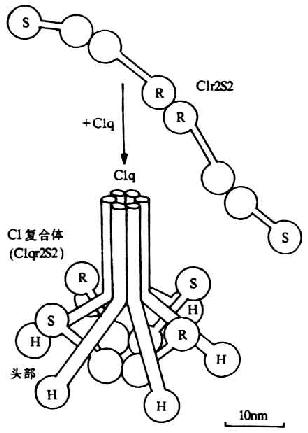
ͼ5-3��C1���ӣ�C1q��C1r��C1s���Ľṹ��ʾ��ͼ��
ĿǰC1r��C1s��cDNA��¡���ѳɹ�����������ȫ�����з���������C1r�Ļ���λ���˵ĵ�12��Ⱦɫ��̱�13-ter�������C1s�Ļ���������
����C4����
C4�Ǿ��伤��;���еڶ�������IJ���ɷ֣�������ԼΪ210kDa���ɦ���90kDa�����£�78kDa�����ã�33kDa����������������������ɣ�ͼ5-4��C4�ķ��ӽṹ��Ϊ���⣬������к���һ���ڰ��װ���Ȱ��������γɵ�����������������N����C1s˿���ᵰ��ø�����õ㡣��C1s��C4�����ľ�����-���������76-77λ���ѽ���γɴ�С���ȵ�����Ƭ�Ρ�СƬ��C4a��8.6kDa���ͷ���Һ���У���Ϊһ���Ĺ������أ����м�̪�����ã����յ��ʴ�ϸ���ͷ��鰷������Ѫ�ܵ�ͨ������ֲ���������֢��������Բ���C3a��C5a��1%�����Ƭ��C4b���`��������������ˮ�⣬����¶��һ�����ɵ��������һ���Ȱ������л��ĸ߶ȷ�Ӧ��������ͨ��ת����Ӧ����C4b�̶���Ĥ�����ϡ���C4bֻ���������������λ��ϣ���߶ȷ�Ӧ�Ե�������Ѹ����H2O��Ӧ�������ȶ������۽�Ϲ��ܵ��Ȼ������ͼ5-7����
һ��C1s˿���ᵰ��ø�����ѽ���C4���ӣ���������C4bֻ��1/10�ܽ�ϵ�Ĥ�����ϣ���������Ҳ��������C2��ϡ�C4b�Ĺ��ܣ�����Ҫ���뾭�伤��;����C3ת��ø��C4b2a����C5ת��ø��C4b2a3b�����γɽ�һ���鵼��������ɷֵļ�����Ӧ�⣬����ͨ����ЧӦϸ��Ĥ�ϵ�CR1��ϴٽ����ɡ����ڲ������Լ������ֹ���߸�����ij������кͲ��������á�������Ϊ��C4����������ʶ��ά���������ȹ���Ҳ�йء�
������C4�Ļ���λ�ڵ�6��Ⱦɫ���HLA-DR��HLA-Bλ���һ�λ�����DNA�ϡ�C4����������C4A*��C4B*�����룬���Ѫ���е�C4����Ҳ���������ͼ�C4A��C4B�������߾��и߶�ͬԴ�ԣ��������������ͬ��ĿǰC4A*��C4B*��cDNA��¡���ѳɹ������������з�����C4A��C4B��B���Ӽ�C2������MHC�ĵڢ�����ӡ�
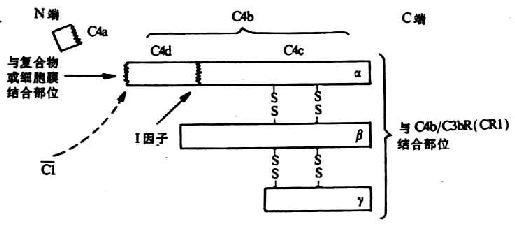
ͼ5-4��C4�������ѽ�Ƭ�Σ�ģʽͼ��
����C2����
C2��������Dz���ĵ�2���ɷ֣����ھ��伤��;���ļ���˳����ȴ��C4�Ժ��C2���ӵ�һ���ṹ��ȫ���������������723��������л���ɵĵ������ǵ��ף�������Լ110kDa��ͼ5-5������C2���ѹ̶���ϸ��Ĥ�����ϵ�C4b���Ϊ������ʱ��C1s˿���ᵰ��ø�ɴ�C2�����ľ�����������ᣨ223-234���䣬��C2�ѽ�Ϊ����Ƭ�Σ���C2a��C2b��C2b��N��223��������л����ɣ�������Ϊ35kDa����ϸ��Ĥ�����ͷ���Һ���У�������ѧ������������C2a��509��������л���ɣ�������Ϊ75kDa�����ǹ��ɾ��伤��;����C3ת��ø��C4b2a����C5ת��ø��C4b2a3b����øԭ���֡�C2a�������Ϻ����ѽ�C3��C5�ĵ���ø���Ե㣬C3ת��ø��C5ת��ø��C3��C5���ѽ⣬������C2a��ø���Ե�������á�
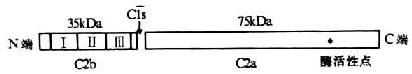
5-5 C2���ӵĽṹ��ģʽͼ��
ע����Ϊ3��SCR
ͨ����C2��cDNA���кͰ��������з������֣�C2��B���Ӽ���нṹ�ϵ�ͬԴ�ԡ�����C2a��BbͬԴ���������ѽ�C3��C5��ø���Ե㣻C2b��BaͬԴ������3��Լ60��������л��Ķ�ͬԴ�ظ����У�shortconsensus repeat,SCR�����ɡ�����C2�Ļ���λ���˵ĵ�6��Ⱦɫ��̱�21��������8kb����
�ġ�C3����
C3������������;���Ļ�ϵ㣬�ڲ���ϵͳ�������������Ŧ���ã���Ϊ���;������Ĺؼ����ӡ�C3�Ħ���������������ɣ�֮���Զ���������ᣬ������Ϊ195kDa�����Ц���Ϊ115kDa������Ϊ75kDa��ͼ5-6��������Ѫ���еĺ�����������������ӣ�ԼΪ0.55-1.2mg/ml��ͬC4����һ����C3���ӵĦ����ڰ��װ���Ȱ���л�֮��Ҳ��һ������������Cys-S-CO-Glu�����˻�״�ṹˮ����γ�һ��ת���Խ�ϵ㣬��Ϊ����C3b��Һ���ϵ������ϵĽṹ������Ҳ��C3�Ի���������ˮ����乹��ı��������B���Ӿ��������ġ��乹��C3b�ķ��ӻ�������C3ת��ø��C3����N��һ��������-˿���������77-78λ���⽫C3�ѽ�ɲ���һ��9kDa��СƬ��C3a���ͷŵ�Һ����ȥ�������ಿ��ΪC3b��ͬʱ��������C3b��`���ھ�N��5kDa�����������ѣ��ڹȰ��������ϳ���һ������ת�����õĸ߶Ȼ���ǵȷ����е��ǻ���R-OH�����۽�ϣ��γ��µ�������ͬ����Ҳ�����ϸ���ϵİ����γ�������-CONH-����������������C3b��ɽ���ڰ�ϸ�����棬������װ�����ͨ������1�������γ��������-SH�����Ӷ����ת���ԡ���Ҫ�ἰ���ǣ������γɵķ�Ӧ�����������ȶ�������60����δ̼ͬˮ�����ת����Ӧ����Ӧ����������ˮ���ӷ�Ӧ�γ��Ȼ����Ӷ�ʹC3bʧȥ���۽�ϵ�������ͼ5-7����һ��ϸ�����涼���㹻�����ࣨ�����ǵ��ס���֬�������ʽ���ڣ��������C3b����ͨ������ת�����ö��̶���ϸ���ϣ������Ļ������ģ���ͨ������ת����Ӧ����ý�ϻ��Ե�C3b����c
4b2a����γɾ���;����C5ת��ø��C4b2a3b��������Bb����γ����;����C3ת��ø��C3bBb����C5ת��ø��C 3bnBbC3bҲ����H���Ӻ�I���ӵ������£���Ϊ���Ե�C3bi������I�����ѽ�ΪC3dg��C3c��ͼ5-8����C3d������C3�ѽ�����ղ��ֻ����ϸ��������ֽ����������£������ѽ�ΪC3d��C3c����������C3eƬ�Σ�������Դ��C3c��
C3�����ѽ�Ƭ�ε�����ѧ���Բ�һ��C3aΪ�������أ���ֱ�������ڷʴ�ϸ����������ϸ����ʹ֮�ͷ��鰷������Ѫ�����ţ�ͨ�����ӣ�ƽ�����������ֲ�ˮ�ס���������Զ��C5a�������⣬C3a������ʹ����ϸ�������ƶ��Դٽ����ɵ��������ã��Լ��������쿹�巴Ӧ���������Զ��¡��Ӧ�����ư�ϸ���ƶ��������ӣ�LIF�����������á�C3b������ѧ���Ԟ�㣬�������������¼������棺��1���������;��������C3ת��ø[��ʼC3ת��ø��C3bB���ͷŴ�C3ת��ø��C3bBb��]���Լ�����;��������C5ת��ø��C4b2a3b��C3bnBb�����γɣ���2�������������;���е��������Ŵ��·����3�������ٽ����ɼ�����ճ�����ã���4���������ߵ��ڣ�����ΪBϸ����ķ������Դ̼��źţ���ΪBϸ��������˿����ԭ�ٽ�Bϸ����ֳ���뿹��Эͬ��ǿADCC���úʹ̼�����ϸ���ͷ�ǰ������E��PGE����Ƕ�뿹ԭ�����帴���������ṹ�У�ʹ���ߵĽ�ϼ����ѴӶ��Dz����Կ��������߸�������ܽ����õȡ�C3bi���дٽ����ɺ��뿹��Эͬ��ǿADCC��Ӧ�����á�C3c��C3dg�ɵ����ƿ�ԭ����˿����ԭ��ͬ�����Ϳ�ԭ�յ���Tϸ����ֳ��C3e��������ϸ�����ࡣ
�˵�C3����λ�ڵ�19��Ⱦɫ�壬�����ֳ�����ͬ����C3S��C3F�������ʮ�����ټ��ͼ������ͣ�����C3F����С��ëϸѪ�������Ͳ���֬����Ӫ�������йء�C3�Ŵ���ȱ���ټ������緢��ȱ�ݣ�����������ŧ�Ժ��������Ծ��ĸ�Ⱦ��

ͼ5-6��C3���ӵĽṹ��ģʽͼ��
�塢C5����
C5���γ�Ĥ���������壨MAC���ĵ�1��������ӡ�C5���Զ���������ӵĦ���������ɣ�������190 kDa�����Ц���Ϊ115kDa������Ϊ75kDa��ͼ5-9����C5��C3��C4�Ľṹ�����ƣ���û������������������N�˵ĵ�74-75λ������һ�������ΪC5ת��ø���õIJ�λ����C5ת��ø�������£�C5����Nĩ���ѽ��һ��������Ϊ11kDa��СƬ��C5a����Һ���У����ಿ��Ϊ110kDa�Ĵ�Ƭ��C5b���Խ����ϸ��Ĥ���档������C5b�ڼ���ʱ�����ܱ�����C6��ϵĹ�����C6�ǹ��۽���γ�һ�ι̵�C5b6�������ͨ����C3b�Ŀ����Խ�϶��̶���ϸ��Ĥ�ϡ���C5b���ɺ���DZ�ڵ�����ѧ���Դ���ʱ��dz��̴٣�����C6�����Ѹ��˥��ΪC5bi��
C5bֻ�γ�MAC����ϸ���ܽ�ЧӦ����C5aȴ���й㷺������ѧ���ԡ��������������¼����棺��1�������������ã�C5a�Ǿ��й����������õIJ����ѽ�Ƭ����������ǿ�Ľ��ʣ���C3aǿ20������C4aǿ2500�������⣬C5a���ɲ������ڷʴ�ϸ���ͷ��鰷����ͨ��ֱ��������Ѫ����Ƥϸ��������Ѫ�ܵ�ͨ�ԡ���2���������ã���Ũ�ȵ�C5a��������ϸ������������ϸ���͵���ϸ�������������ɴ̼���Щϸ������Ũ�ȶ����ƶ���ֵ��ע����ǣ���Ѫ������øN��C5a C�˾�����л����γɵ�ȥ��C5a��ɥʧ��ʹ�ʴ�ϸ�������鰷�����������Ծ��н�ǿ���������ԣ��Dz���������������õ���Ҫ���ء���3���ٴ�л���ã���Ũ�ȵ�C5a�ɴ̼�������ϸ���͵���ϸ����������л�������cGMP��ˮƽ�������ڴٽ���ø����ϸ��Ĥ���ںϣ��ͷ���ø��ø�����⣬C5a���ɴ̼�������ϸ��ճ������ǿ��������������4�����ߵ������ã����������о�������C5a������Ӧ����������ǿ���ã�����յ�����ϸ������IL-1��IL-6��IL-8��TNF-����ϸ�����ӣ��ٽ���ԭ��ͬ�����忹ԭ�յ���Tϸ����ֳ��Bϸ����������ȡ�C5a����������ѧ���Ե�������ǿ����ķ������ܣ������䵼�µ���֢��ӦҲ����ɶԻ�������ˡ�������C5�Ļ���λ�ڵ�9��Ⱦɫ�峤��32-34����
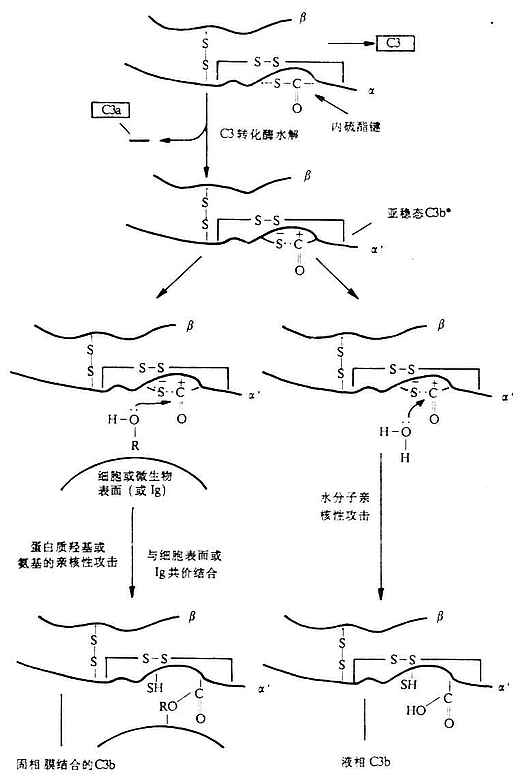
ͼ5-7��C3b��������Ӧ��ͼ�⣩
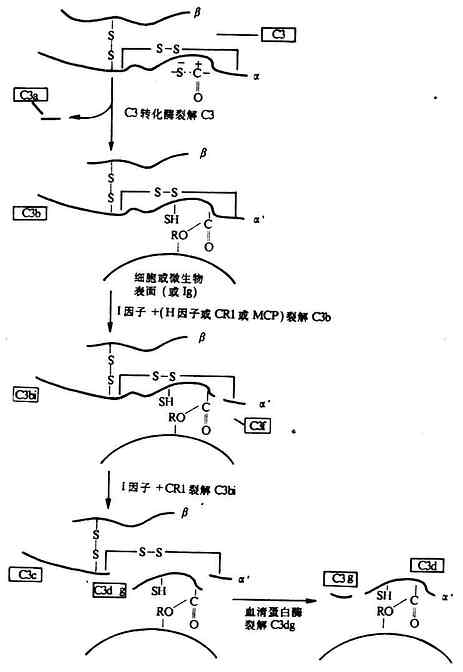
ͼ5-8��C3�����ѽ�Ƭ�εIJ�����ͼ�⣩
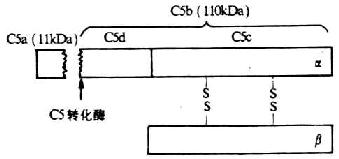
ͼ5-9��C5���ӵĽṹ��ģʽͼ��
����C6��C7
C6��C7����������֮������Ϊ�����ǵ��ף��ҷ�����Ҳ����ֱ�Ϊ128kDa��121kDa������C6��C7���ӵĻ�������ɹ�ͬ����������������C6��C7�ڰ�����ˮƽ����33.5%��ͬԴ�ԡ�������������C6�Ľṹ�����ܽ����˽�������о�����cDNA�����Ƶ�����C6��ȫ������������913�������л���ǰ�滹��21�����ذ�����л���ɵ��ź��ģ���̼ˮ����ĺ���Ϊ4-6%���������ĵ�303λ��834λ������л���������Ϊ�����춬�������ӵ��ǻ�����λ��C6�л����д����İ��װ���л�������Ϊ64�����������ڶ������İ���ĩ�˺��Ȼ�ĩ�˲��֣����а���ĩ�˵�λ���ɰ��װ���л���ռ�ݡ�
C6��C7�ж����е��ܶ�֬���ף�LDL������ṹ������EGFǰ��ṹ����������Ѫ���е��ף�TSP-1���ṹ�������SCR�ṹ�����������з�ʽ��ͬ��Ӧ����ֽ��ϵ�C6Ƭ�ν����о�������C6��C5b�Ľ�ϲ�λΪ��2��SCR��2�������ӽṹ������FIMs������ɵĴ�СΪ34kDa���Ȼ�ĩ�˵�Ƭ�Ρ���C6��C7������У����߾����ӵ��ѽ⣬�Ʋ��������������ӹ��͵ĸı����Ϊ���н�ϻ��Ե���ʽ��C6��C5b�Էǹ�����ʽ����γɵ�C5b6�����������ɵ���C3b�ʿ����Խ�ϣ��Ҿ�����ˮ�Բ��ܲ���Ĥ�ڡ���һ����C7��ϣ���������ˮ-��ˮ����ת����ͬʱ��������̬Ĥ��ϲ�λ��������c
5b67������C3b���Ų�λת����Ĥ���棬Ȼ��ͨ����������C7����ˮ�Խ����̶���Ĥ֬��˫���С���ˮ���ı�¶ϵ����C5b67������Ĺ���仯���¡�������������̬C5b67���������100����������䣬�粻��ʱͬĤ��ϣ��ֿ���������ʹ��С���۵���ʧȥĤ��ϻ��ԡ����⣬����Һ����ϵ�Ĥ�ϵ�C5b67������������оۺ϶�ɥʧ��鵼����ϸ�����ԣ����Ծ����������á�C6��C7���ܻ��д����ܰ�ϸ��ĸϸ���������ã����ڵ������ܰ�ϸ����Ӧ��MLR���У����뿹C6��C7�Ŀ���Fab�������ܰ�ϸ����ֳ��
����C6��C7�͵Ļ���λ���˵ĵ�5��Ⱦɫ��������������C6��C7�������Ŵ���̬�ԣ�����C6��������λ����C6A��C6B����ȷ����������Ⱥ��C6B�Ļ���Ƶ�ʽϸߡ�C6��C7��cDNA�ѿ�¡�ɹ�������������C8��C9����һ����ͬԴ�ԡ�
�ߡ�C8����
C8���ɦ����¡�������������ɵ��������ǵ��ף�������Ϊ155kDa�����Ц����ͦ�����Ϊ64kDa,����Ϊ22kDa�������ͦ������Զ�������۽�ϣ����������������Ϊ�ǹ��ۼ���ϣ�ͼ5-10����C8������Ҳ����TSP-1��LDL����ṹ��������C8����C8�¶����������루157-501��������л��䣩�����Ʋ������װ���л���Ϊ��ϸ������Tϸ����NKϸ�������Ĵ����ף�perforin����ͬԴ�ԵĽṹ�������ڦ��ͦ����к��м��߱�ʹ����ˮ�����ᡣ�����ֲ���C8���ӵı��棬����C5b��������Ǽ��Եģ������и߶������ԡ�C8��C5b-7�Ľ�ϲ�λΪ���������C8��c 5b-7��Ϻ�ͨ��C8���ӵĹ���仯��ʹ���������Ĥ֬��˫��������У��γ�ֱ��Լ1.6nm�Ŀ�Ĥ������ʹϸ��ͬ�����ӻ��������������ᵼ��ϸ���ܽ⡣C5b-8�������ܴ�ʹC9�ľۺϵ������в�����������ǽ�����C9�ۺϵĻ�����¡������о�������C9��ͨ��C8��ͬc 5b-8��ϵģ���C9����ͬC5b-7���ϣ�����ͬC5b-8�Ľ����ɱ���C8�Ŀ��������ơ�
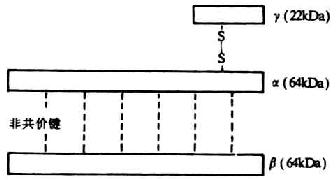
ͼ5-10��C8���ӵĽṹ��ģʽͼ��
C8�Ļ���λ�ϸ��ӣ����б�������ͦ����Ļ���C8A��C8B��λ���˵ĵ�1��Ⱦɫ�壬����������Ļ���C8G��λ�ڵ�9��Ⱦɫ��ij��ۡ�Ŀǰ�Ѷ�C8������cDNA��¡�ɹ������������з�������������C9���и߶ȵ�ͬԴ�ԣ����Ҷ��߾����зḻ�İ��װ����ĤǶ������C8�Ħ����ͦ������Ŵ���Ҳ�ʸ߶ȶ�̬�ԣ�����Լ��33%�İ�����������ͬ������C7��C9��Լ25%��ͬ
�ˡ�C9����
C9���γ�Ĥ���������壨MAC�����������ӣ�Ϊһ�����ǵ��ף�������79kDa������cDNA�Ƶ��İ��������з������֣�C9Ϊһ���Է��ӡ�C��37kDa����ˮ��������ɳ�C9b��N��34kDa����ˮ��������ɳ�C9a���C9�����Ȼ��˲���Ƕ��ϸ��Ĥ��֬��˫���С���N����Ϊ��c 5b-8���ϵĽṹ��C9�����Է��ۺϵ����ã����ۺϺ�������37������3�������ɣ�����C >5b-8�Ĵ��£�10�����ڼ�����ɡ���12-16��C9���Ӿۺ��γɵĶ����C9�����γ��ھ�10nm���ں�2nm���пմ�Ĥ��Ƕ��Ĥ�ڣ�ͼ5-11��������������������ˮ������л���̼ˮ������ɣ�����˫��֬�Ӵ��Ĺܱ�����������ˮ������л�������ϸ�����������©�����տɵ���ϸ���ܽ��ƻ���C9���ӵĶ�������C8����C8�½ṹ�������ƣ�Ҳ����TSP-1��LDL����ǰ��ṹ�������봩����ͬԴ�Ľṹ����������C9�ʹ������ڽṹ�����Ͼ��dz����ƣ��Ʋ���߿��ܾ��й�ͬ�����������C9�Ļ���λ�ڵ�5��Ⱦɫ���ϣ�ĩ����C9�ж�̬�ԡ�
�š�B����
B���ӣ�factor B,Bf���������;���е���Ҫ�ɷ֣���Blum��1959�����ȷ��֡�B����Ϊ��733��������л���ɵĵ����ǵ��ף��Ǻ���Լ7%����������93kDa��������Щ��������ػ��۵��γ�������С����Ƶ�������������1��ΪBa����������������״ΪBb��Bb�п���N�˵�һ����������ͬC3b��ϣ���һ�������������Ǵ�����ͼ5-12������Mg2+���ڵ�����£�B���ӿ���C3b����γ�C3bB����Ѫ���е�D�����ѽ�Ϊ������Ϊ33kDa��Ba��63kDa��Bb����Ƭ�Ρ�����3����C3b����γ����;����C3ת��ø��c3bBb����C5ת��ø��C3bnBb��������ø�е�Bb������˿���ᵰ��ø���ԣ����ѽ�C3��C5�Ļ��Բ�λ����C 3bBb��C 3bnBb�����ȶ�����˥��ʧȥ���ԡ�

ͼ5-11��Ĥ���������壨MAC���Ľṹ��ģʽͼ��
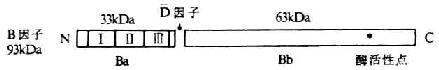
ͼ5-12��B���ӵĽṹ��ģʽͼ��
ע��Ϊ3��SCR
�����о����֣�B���ӵ������ѽ�Ƭ�λ��������ߵ������á�����Bb�ܴٽ������ɫ�������Cowan I�꣨SAC���̼����Bϸ����ֳ����Ba���Bϸ���������ӣ�BCGF���յ��Ľ���״̬��Bϸ����ֳ�������������ã��ҳ�Ũ��������ϵ��B���Ӻ�C2�����ڲ��峬����ij�Ա�����ߵı�������������������B���ӵĻ���λ���˵ĵ�6��Ⱦɫ��̱�21��������6kb����18�������ӡ�
ʮ ��D����
D�������������;���������Ҫ�ɷ֣�Ϊ��222��������л���ɵĵ���˿���ᵰ��ø����������25kDa��D������Ѫ���е�Ũ�Ⱥܵͣ�1-2��g/ml������Ҫ�Ի��ʽ�����ڡ������ܻ���һ����øԭ��ʽ�����ڵ���239��������л���ɵ�D���ӡ����л��Ե�D���ӣ�D�������ڵ�234-235λ�ľ�����-�����������B�����ѽ�ΪBa��Bb����Ƭ�Σ��Ӷ��������;���ļ������Ӧ��D���ӵIJ���cDNA�ѿ�¡�ɹ��������������з���������������������˿���ᵰ��ø�����ȵ���ø���ӵ���ø������ø�����Ե���ø������ͬԴ�ԡ�
ʮһ��P����
P�����ֳƱ����أ�properdin���������;���г�C3�������ȷ��ֵ�һ��Ѫ�����ס�����̽����P�����Ծۺ�����ʽ�����ڣ��������壨54%���������壨26%�����ľ��壨20%�����У���������Ե�˳������Ϊ���ľ���>������>�����塣P����Ϊ��4����ͬ����������������55kDa����ɵ��ľ�����ӣ������Էǹ��ۼ������ӣ�������Ϊ220kDa��P���ӵ�����ѧ�������Ը�������c 3bBb��C 3bnBb���ϣ���Ϻ�ͨ����������ı���ӹ�C3b��Bb��Ľ�������Ӷ���ʹ���˥����2�����ӳ���26���ӡ����⣬P���ӻ��ɷ��H���ӵ��������ã�����������������ø���ȶ��Լ����ԣ������ڴٽ����;��������Ӧ�ļ������С���ˣ�P����ʵ���������;���е�һ����Ҫ�������ڷ��ӡ����䳣��Ϊc 3bBb��C3bnBb�������е���ɳɷ�֮һ���ʽ�����Ϊ����ϵͳ�Ĺ��гɷ��ڴ�һ�����������⣬��Ĥ��������С�����ײ���Ѫ���з�����һ��C3�������ӣ�C3nephritic factor,C3NeF��ʵ��ΪC 3bBb���������壬Ҳ����C3bBb��϶�����c 3bnBb���ȶ��ԣ�ʹ���˥�ڴ���10-30����
��������14�ֲ�����е������Լ�������ѧ���Լ���5-1��
��5-1��������гɷֵ����Լ�����ѧ����
| ����ɷ� | Ѫ��Ũ�ȣ���g/ml�� | ��������kDa�� | �ǵ�λ����������������kDa�� | ������� | ����ѧ���� |
| C1q | 75 | 410 | A:24��6�� | �� | ʶ��IgG��IgM Fc�IJ����ϵ� |
| �� | �� | �� | B��23��6�� | �� | �� |
| �� | �� | �� | C��22��6�� | �� | �� |
| C1r | 50 | 85 | 1�� | C1r | ˿���ᵰ��ø���ѽ�C1s |
| C1s | 50 | 85 | 1�� | C1s | ˿���ᵰ��ø���ѽ�C4��C2 |
| C4 | 200-500 | 210 | ����90 | C4a | �������������� |
| �� | �� | �� | �£�78 | �� | �� |
| �� | �� | �� | �ã�33 | C4b | ���CP�е�C3��C5ת��ø���ٽ����ɡ���ֹIC�������кͲ���������ʶ��ά�������ȶ��� |
| C2 | 20 | 110 | 1�� | C2b | ���� |
| �� | �� | �� | �� | C2a | ˿���ᵰ��ø�����CP�е�C3��C5ת��ø |
| C3 | 550-1200 | 195 | ����110 | C3a | �������ء��������õ� |
| �� | �� | �� | �£�85 | C3b | ���CP��AP�е�C3��C5ת��ø�����������ɡ�����ճ�������ߵ������� |
| C5 | 70 | 190 | ����115 | C5a | ǿ�Ĺ����������á��������á� |
| �� | �� | �� | �£�75 | �� | �ٴ�л���ü����ߵ������� |
| �� | �� | �� | �� | C5b | �γ�C5b67����������������� |
| C6 | 60 | 128 | 1�� | �� | ���MAC�������ܰ�ϸ��ĸϸ���� |
| C7 | 60 | 121 | 1�� | �� | ���MAC�ijɷ� |
| C8 | 60 | 155 | ����64 | �� | ���MAC�ijɷ֣��ٽ�C9�ۺ� |
| �� | �� | �� | �£�64 | �� | �� |
| �� | �� | �� | �ã�22 | �� | �� |
| C9 | 60 | 79 | 1�� | �� | ���MAC���ۺ��γɿ�Ĥ�� |
| B���� | 200 | 93 | 1�� | Ba | �����˵�Bϸ����ֳ |
| �� | �� | �� | �� | Bb | ˿���ᵰ��ø�����AP�е�C3��C5ת��ø |
| D���� | 1��2 | 25 | 1�� | D | ˿���ᵰ��ø���ѽ�B���� |
| P���� | 25 | 220 | 4������55 | �� | �ȶ�AP�е�C3��C5ת��ø |
ע��CP����伤��;����AP���������;����MAC��Ĥ����������