�ڶ��ڡ���ϸ���ֻ���ԭ��Ӧ��
CD��ԭ������Ӧ�ĵ���¡�����ڻ������ٴ�����ѧ�о����ѵõ��㷺��Ӧ�á��ڻ�������ѧ�о��У�CD��ҪӦ���ڣ���1��CD��ԭ�Ļ����¡����CD��ԭ��������ķ��֣���2��CD��ԭ�ṹ�빦�ܹ�ϵ����3��ϸ������;����Ĥ�źŵĴ�������4��ϸ���ֻ������еĵ��أ���5��ϸ����Ⱥ�Ĺ��ܡ����ٴ�����ѧ�о��У�CD����¡��������ڣ���1���������߹��ܵļ�⣻��2����Ѫ�����ܰ������߷��ͣ���3�����߶��������������ơ�������ֲ�Լ���ֲ�ųⷴӦ�ķ��Σ���4���������ߵ������ơ��й������߹�����ص�CD���ӹ����ڱ�1-4���й���Tϸ��������ӡ�Bϸ����������Լ�NKϸ���ı����Dzμ������¡���CD�йص�Ig�����塢ճ�����ӡ��������塢ϸ����������ȷֱ��ڱ�����й��½��м��Խ��ܡ�
��1-4�������߹����йص�CD
| ���߹��� | CD |
| ϸ������ TCR��CD3��CD4��CD8 mIg��mb-1/Ig����CD79a)��B29/Ig��/(CD79b) CR��CR1(CD35)��CR2(CD21)��CD3(CD11b/CD18)��CR4(CD11c/CD18)��C5aR(CD88)��FcR��Fc��RI(CD64)��Fc��R��(CD32)��Fc��R��(CD16)��Fc��R��(CD23)Fc��R(CD89) ϸ������������IL-2R��(CD25)��M-CSFR(CD115)��GM-CSFR(CDw116)��SCFR(CD117)����IFN-��R(CDw119)��TNF-��R(CD120)��IL-1R(CDw121)��Il -2R��(CD122)�� IL-4R(CDw124)��IL-6R(CD126)��IL-7R(CDw127)��IL-8R(CDw128)�� gp130(CDw130) ϸ���䡢ϸ�������ʶ�� ��ϸ��ճ������-��Ƥϸ��ճ�����ӣ� LFA-1��CD11a/CD18)-ICAM-1(CD54)��ICAM-2(CD102) Mac-1(CD11b/CD18)-ICAM-1(CD54) VLA-4(CD49d/CD29)-VACM-1(CD106) L-selectin(CD62L)-E-selectin(CD62E)��P-selectin(CD62P) CD15-E-selectin(CD62E)��P-selectin(CD62P) �ܰ�ϸ����λ����-Ѫ����Ƥϸ����ַ���� L-selectin(CD62L)-PNAd CLA-E-selectin(CD62E) LFA-1(CD11a/CD18)-ICAM-1(CD54)��ICAM-2(CD102) VLA-4(CD49d/CD29)-VCAM-1(CD106) CD44-MAd LPAM-2(CD49d/��7��-MAd��VCAM-1(CD106�� ��ϸ��ճ������-ϸ������ʣ� VLA-1(CD49a/CD29)-CA��LM VLA-2(CD49b/CD29)-CA��LM VLA-3(CD49c/CD29)-FN��LM��CA VLA-4(CD49d/CD29)-FN VLA-5(CD49e/CD29)-FN VLA-6(CD49f/CD29)-LM ��7��1��-/CD29��-LM VNR-��1��CD51/CD29��-VN��FN Mac-1(CD11b/CD18)-FB P150,95(CD11c/CD18)-FB GP��b��a(CD41/CD61)-FB��FN��vWF��TSP VNR��CD51/CD61��-VN��FB��vWF��FN��CA��TSP ��6��4��CD49f/CD104)-LM VNR-��5(CD51/-)-VN CD51/ ��6-FN CD49d/��7-FN GPIb-��/IX��CD42b/CD42a)-vWF ����ϸ�����ʶ�� CD22-CD45RO CD2-LFA-3(CD58) CD4-MHC������� CD5-CD72 CD 8-MHC������� LFA-1��CD11a/CD18)-ICAM-1(CD54)��ICAM-2(CD102) CD28-B7/BB1��CD80�� CD27-CD70 �����ϸ������ Tϸ����CD2��Tϸ����·����;������CD3���ź�ת������CD4��CD5��CD8��CD28����CD43��CD44��VLA-4��CD49d/CD29)��CDw90 Bϸ����CD19������G0��C1������Ig���ڣ���CD20������ϸ�����ڣ���CD21���Bϸ��������CD22��Ca2+���ߣ��ٽ�G0��G1����CD23(Bϸ���ֻ�����CD40��Bϸ������������CD72��CD73��G0��G1����CD80��Bϸ����� ����ϸ����CD14������ϸ����������������CDw65��������ϸ����� NK��CD16��NK�����CD2��CD3 ����ϵ��CD69����յ����ӣ�AIM�� ��ϸ��Ĥ����ø�й� CD10��������������ø����CD13������ø����CD26����������ø������ CD45���Ұ���������ø���� CD73��5`����������ø����CDw75������ø���ԣ� �벡�������й� CD21��EB����R����CD4��HIVR����CD54���Dz���R����CD46����������壩 | |
һ����Tϸ��ʶ��ճ������йص�CD����
Tϸ����һ����Ҫ��������ϸ������ֱ�ӽ鵼ϸ�����߹����⣬�Ի�������Ӧ��ĵ�����ؼ����á�T�ܰ�ϸ��������ʶ����ЧӦ���ܵķ��ӣ�������������ԭ��˿��ԭ�Ͷ���ϸ������������أ�����������Tϸ���֮�䡢Tϸ���뿹ԭ���ϸ����APC��֮���Լ�Tϸ�����ϸ��֮���ֱ�ӽӴ���T�ܰ�ϸ��ʶ��ԭ��������Tϸ�����壨t cell receptor,TCR)��CD3����ɵĸ����TCR/CD3������ʶ������л������ڿ�ԭ�������Ե�����ϸ��������ӵĸ�������Щ�������ӣ�accessory molecules)��Ҫ����CD4��CD8��MHC ������ӡ�������ӣ�LFA-1��CD11a/CD18)��CD49d��e��f/CD29(VLA-4��VLA-5��VLA-6����CD28��CD44��CD45��ICAM-1��CD54����LFA-2��CD2����LFA-3��CD58���ȡ�
�й�MHC���ࡢ������ӵĽṹ�����ڵ����¡�MHC�����ٴ�Ӧ�á������ۡ�VLA-4��VLA-5��LFA-6��LFA-1��ICAM-1��CD44���ڶ��¡�ճ�����ӡ����й�CD45�ڵڰ��¡������׳����塱�в�����
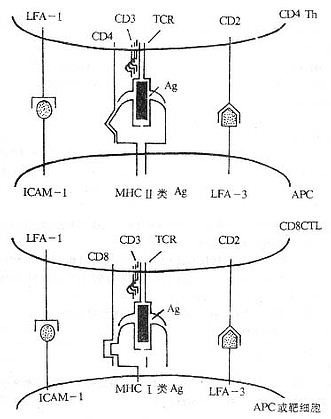
ͼ1-1 ����Tϸ����ϸ��ʶ��ķ��ӣ�ģʽͼ��
Tϸ������ĸ��������������ص㣺
��1��������Tϸ���ϵĸ������ӿ�������������APC���ϸ���ϵ�ijЩ���ӣ����壩���ϣ���LFA-1��CD2�ɷֱ���ICAM-1��LFA-3��ϡ�
��2���������ӱ��������ж�̬�ԣ���һ���������и��������Tϸ����ijһ�ָ������ӵĽṹ����������ͬ�ġ�
��3���������ӿɼ�ǿTϸ����APC���ϸ����ϵij̶ȡ�
��4�����ศ�����Ӿ���ת���źŵĹ��ܣ���CD2��CD4��CD8�ȷ��ӡ�
��5����Щ����������CD2��CD4��CD8��CD28��Thy-1�������Ļ�������Ig�����壻��Щ����������LFA-1��VLA-4��VLA-5��VLA-6�ȱ���Ļ�������integrin �����塣
��6��Tϸ��Ĥ���渨��������ΪĤ������Ҫ�ı���ѱ�Ӧ�����ٴ�����Ϻ����ơ�
��7��ϸ�����ӿɵ��ڸ������ӵı���Ӷ��ı�ϸ����ճ��������������ϸ���������ߵ������õ�һ����Ҫ���档
��һ��Tϸ������
Tϸ�����壨T cell receptor,TCR��Ti����T�ܰ�ϸ������ʶ��������ԭ������MHc ���ԭ������ԭ������������壬��ͬ��������ֲ��TCRҲʶ���ķ��ѵ�MHC��ԭ��Ŀǰ�Ѿ�֤ʵ��TCR��ϸ��������CD3���н����һ�����TCR/CD3�����TCRʶ��ԭ��̼��ź���ͨ��CD3���Ӵ��ݵġ�
1.Tϸ����������ͺͽṹ��TCR�еĶ������������Եġ����ݿ�ԭ�ṹ�ͱ������ͬ���ѷ����Ц����¡��úͦ����ֶ�����������TCR�������Ľṹ����Ǵӷ���TCR������cDNA��������¡��genomicclones)����������TCR�������Ļ������������������Ա������TCR������������40��60kDa֮�䡣����TCR����Դ˫�����ɵIJ�ͬ��TCR�ɷ�Ϊ��TCR���º�TCR�æ��������͡�
��1��TCR���£�CD����TCR����Tϸ����ʶ�����MHC���ԭ��ͬ�����忹ԭ��������MHC���ԭ��ӹ���ԭ�ĸ�����CD8����TCr ����Tϸ�����ʶ����MHC���ԭ������MHC ���ԭ��ӹ���ԭ�ĸ�����TCR����������40��50kDa�������ǵ��ף�����40��50kDa�����������ǵ��ס����ͦ�������һ���ɱ�����V������һ���㶨����C������ɣ���Ig��V����C����С���ƣ����������׳������Ա��TCR����������V��Լ��102��109�������ᣬ��V���������������װ����γ����ڶ���������Լ��50��60������л��Ļ��ģ�����IgV���ṹ�������ƣ���������ʶ��������ԭ�Ľṹ��TCR����������C��Լ��138��179�������ᣬÿ��C���γ������ڶ�������ӵĻ��ġ����������������ģ�connectingpeptide)�γ�������������Ĥ��Լ��20��24���������,������Ĥ�����д������1���������1��������л�,������Ĥ������1���������������л�,��Щ������İ�������CD3�á��ĺͦ�����Ĥ��������ĹȰ����/���춬�����γ����ţ��ȶ�TCR/CD3������ṹ������CD3������Ϣ�йء�����������������ֻ��5��12�����᳤��β����ͼ1-2����

ͼ1-2 TCR������Դ˫��ģ
��2��TCR �æģ�TCR�úͦ���������һ��Ig����V����C���������ġ���ˮ�Ĵ�Ĥ���Լ�һ���̵İ�����β�����������������γ�����Ķ�������úͦ����Ĵ�Ĥ��������1��������������ᣬ�����������1��������ľ����ᣬ��Щ������İ�������CD3�á��ĺͦ�����Ĥ����������춬�����Ȱ����γ����š��ڰ�����ˮƽ�Ϸ�����TCR���������ͬԴ�Խϸߣ���TCR���������ͬԴ�Խϸߡ�������TCR�æ��ж���������ͷǹ�������������ʽ������С��ֻ���ֶ����������TCR�æ���ʽ���˦���������Ϊ36��55kKa������Ϊ40��60kDa���á������ķ�������Сȡ���ڶ��ĹǼܵij��Ⱥ��ǻ����ij̶ȡ�
�й�TCR�����¡��á���������Ľṹ�����ż������¡������׳����塱
2.��������TCR Tϸ���ıȽϡ�TCR������TCR�æIJ����������������Ľṹ��ͬ�����Ҿ�����������������Tϸ���ķֲ������͡������Լ�����Ҳ�в�𣨱�1-5����
��1-5��TCR������TCR�æ�ϸ�����ԵıȽ�
| ���� | TCR���� | TCR�æ� |
| |
| �֡��� | PBL | 60��70% | 0.5��15% |
|
| �������� | �� | С����ͻ״��Ƥϸ����DEC����С��ճĤ��Ƥ���ܰ�ϸ����IEL�� |
| |
| ������ | CD4+CD8- | 60��65% | <1% |
|
| CD4-CD8+ | 35% | 20��50% |
| |
| CD4-CD8- | <1% | 50��80% |
| |
| CD2 | 100% | 100% |
| |
| CD5 | >95% | -��+������ |
| |
| ������ | ���ٺ��� | �������ڣ�����TCR���£� |
| |
| ������ | ��1��ʶ��MHC��ӹ����� ���ĵĸ����� | ��1��ʶ��������ԭ��MHC�����������ۣ�ʶ�����˷���ؿ�����MHC���ԭ���ƣ�С��TCR�æĶԺϳ��ķ�Ӧ��Qa�������ơ� |
| |
| ��2��ʶ�����MHC��MHC���ԭ��ط��ӣ�TLa��CD1�� | ||||
| ��2��ʶ�����MHC | �� | |||
| ��3������Щϸ�����ӣ�IL-2��IL-4��IL-5��GM-CSF��IFN-�ã� |
| |||
| ��3�����������ܰ����� | ��4��ijЩTCR�æ�ϸ����ɱ�˰�ϸ�� |
| ||
| ��5������TCR���µķ����� | ||||
| ��4��ɱ�˲�����Ⱦ�ȰС�ϸ�� | �� | |||
| ��6����Ƥ���ϣ���Գ����ء���֦�˾����ݿ˵��ȣ� |
| |||
ע����ͻ״��Ƥϸ��dendriticepidermal cell, DEC
��Ƥ���ܰ�ϸ��intraepithelial lymphocyte��IEL
����������Ѫ�У�CD4-CD8+��CD4+CD8-��CD4+CD8+��CD4-CD8-���ֱ��Ͳ�ͬ��Tϸ���ֱ�ռTϸ��������25%��70%��1%��4%���ң�����ǰ���ֱ���TCR������ҪΪTCR���£���CD4-CD8-Tϸ����ҪΪTCR�æġ����¼����ɼ�����Ѫ��ֲ�TCR�æ�ϸ��������������ߣ���1����֢��������ȱ�ݡ������ɱ�������ȱ�ݡ�Wiskott-Aldrich�ۺ�����Di-George�ۺ�������Ѫ�����߹�����ֲ�Ȳ�������Ѫ��TCR�æ�ϸ���ٷ������ӣ���2����������Tϸ����Ѫ����Tϸ�������ܰ�������ΪTCR�æ�Tϸ���������Ա䣻��3�������ܰ�ϸ����Ѫ����������ܰ�ϸ����LGL����Ѫ������PBMC��TCR�æ�Tϸ���ٷ������ӣ���4������ֲ�������췴Ӧ��������Ѫ��TCR�æ�ϸ�����ӣ���5�����ʪ�Թؽ����߹ؽ�ǻ��Һ��TCR�æ�����ϸ������Ҫ��������Ѫ��TCR�æ�ϸ���������Ʋ�TCR�æĿ��ܲ���ֲ���֢�ķ�������6������˸˾����ߺ�ľֲ��ܰ�����Һ��TCR�æ�Tϸ���������ӣ�����ڲ������кܸ߱�����TCR�æ�Tϸ������ʾTCR�æ�Tϸ���Է�֦�˾����µĸ�Ⱦ����������Ҫ���ã���7��HIV��EBV��Ⱦʱ����ѪTCR�æ�Tϸ���������ӡ�
[����ԭ]�еĿ�ԭ������APC�����͵ݳʿ�ֱ�Ӽ���CD4����Tϸ����Ϊ����ԭ��superantigen,SAg),���������·���ԭ�����á�SAg��Tϸ���ļ����ȡһ�ֶ��صķ�ʽ��������һ�˺�TCR������V��������ϣ���һ�˺�APC����MHC����������ϡ����SAg����������������ϸ��������TCR������CD4+Tϸ���ͱ���MHC���ԭ�ĸ���ϸ������Դ�Ե�SAg��Ҫ�����Ѿ����������֧ԭ�����������Ķ��أ��������������������A��Staphylococcus enterotoxin A,SEA���о�����࣬SEA����ȡ���ض�TCR����Ƭ��V��6.9��V��22��SEA��һ����HLA-DR���Ӧ�1�ṹ��Ħ��������ϡ�
SAg����CD4+Tϸ��ʹ֮�ͷ�IL-2��IFN��TNF��ϸ�����ӣ��յ�CTL�ֻ�ΪЧӦϸ������ɱ�˶�NK��LAK�߶ȵֿ��İ�Ѫ��ϸ����
������CD3��T3��Leu4��
CD3���ӷֲ��ڳ���T�ܰ�ϸ�����棬�����ɦá��ġ��š��ơ���5�ֶ�������ɣ���Tϸ����ԭ����ǹ������ӣ�ͼ1-3����CD3����¡������յ�CD3���ĺ�TCR��ñ�γɣ�co-capping)�����յ�T�ܰ�ϸ�����TCRʶ��������ԭ������MHC�����γɵĸ����CD3�����źŵĴ��ݾ�����Ҫ���á�
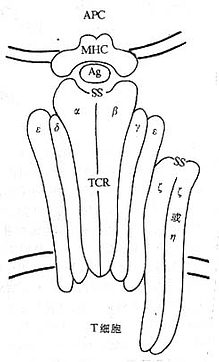
ͼ1-3 TCR/CD3�ṹģʽͼ
Tϸ�������ٷ��������У�CD3�á��ĺͦŻ���ı���Ҫ����TCR������������ı��CD3�á��ĺͦŻ������ͨ�������������γɺ��Ľṹ������֯�������˺��Ľṹ��TCR������Դ˫���γɸ������ת�Ƶ��߶������壬����N���ӵ��ǻ�������-��ͬԴ˫����TCR����/CD3�æĦŦŽ�Ϻ����һ�������ĸ�����TCR����/CD3�æĦŦŦƦƣ���¦����ΪTCR����/CD3�æĦŦŦƦǣ����������һ��������Ϊ28kDa�Ħ�����Tϸ��������ص��ף�t cell receptor associated protein,TRAP),���ܾ��п���TCR/CD3����������֯����װ���ת�ƵĹ��ܣ���ȷ�еĻ����в����ˡ�������������ϸ��Ĥ���档

ͼ1-4 TCR/CD3������ģʽͼ
1.CD3�á��ĺͦ�����CD3�á��ĺͦ��������и߶ȵ�ͬԴ�ԣ�������λ�ڵ�11��Ⱦɫ�壬С��9��Ⱦɫ�壬���������Ļ�����ܴ�һ�����Ȼ���ͨ�������ƶ�����CD3�á��ĺͦ�����ϸ��Ĥ�ⶼ��һ��Ig���ṹ��C2���������������׳����壬�������ڶ�̬�Ի�ɱ��ԣ���˲�ֱ�Ӳ���������ʶ��ԭ���á��ĺͦ����Ĵ�Ĥ���ֺ��д�����Ȱ����/���춬����л�������TCR����������Ĥ���д����������ᡢ�������Լ��¡�������Ĥ���е����������������Ҫ���á��á��ĺͦ����������ֺ�44��81������л����ṩ�˰���Ϣ������ϸ���ڵ�����������������Ϊ25��28kKa����2���ǻ����㣬������89��������л�Ϊ��ˮ�ԣ���ɰ�Ĥ��������Ĥ����27��������л�����������44������л���������113λ˿����л����������ữλ�㡣����������Ϊ20kDa������2���ǻ����㣬������126λ˿������������ữλ�㡣CD3���������ܷ������ԵػTϸ�����ٽ�Tϸ����˿���ѡ�����������Ϊ20kDa����������104��ˮ������İ�Ĥ��������Ĥ��26��������л�����������81��������л���Ŀǰ���Ʊ��ĵ���¡�����д������CD3������
2.CD3�ƺͦǡ��ƣ�zeta)�ͦǣ�eta)���ṹ���ƣ�����CD3�á��ĺͦ�����ͬԴ�ԡ��ƺͦ����������ֱ�Ϊ16kDa��21kDa�����ǵİ�Ĥ���Լ���Ĥ���ͽṹ���ƣ����а�������ͬ����Ĥ�����̣ܶ�ֻ��9��������л������а��װ��ᣬ�Ʀ�֮���Ʀ�֮���γɶ�������ƺͦ�����Ĥ���ָ���һ����������춬���ᡣ�ƺͦ������������ֱ���113����155��������л������ж���Ұ������ữ��λ�㡣����о�֤ʵ��CD3����������NKϸ����Fc���������������⣬������Fc��R����ǵ�λ�кܸߵ�ͬԴ�ԡ�
CD3�á��ĺͦ����ǵ������������Ԧ�-��ͬԴ˫�������80��90%Tϸ���У���10��20%Tϸ�����Ԧ�-����Դ˫����ڡ���������TCR/CD3������������ʽ��TCR����/CD3�æĦŦŦƦơ�
�����⣬��CD3McAb�ɴٽ�Tϸ������IL-2R������IL-2��TNF-����TNE-�¡�IFN-�ú�IL-4�ȶ���ϸ�����ӣ��յ���MHC���Ƶ�ϸ�������ã���ǿTϸ����LAK��NKϸ����ɱ���������á�
1986������FDA����Ӧ��С��CD3 McAb���Ƽ�������ֲ�ųⷴӦ��CD3McAb�����ġ�����ֲ�ųⷴӦ����ɢ����ٴ����飬������Ͷ���г���
������CD4
CD4��CD8���ӷֱ���MHC����͢��ԭ��ϣ���������ǿT�ܰ�ϸ����APC���ϸ����ϵij̶ȣ�������̼��źŵĴ����йء�CD4����ϸ����MHC���ԭ���Ƶ�ϸ��Ⱥ��CD8����ϸ����MHC���ԭ���Ƶ�ϸ��Ⱥ���й�CD4��CD8��ԭ������ϸ���ֻ������еı仯�Լ�CD4��CD8Tϸ����Ⱥ�������¡��ܰ�ϸ��Ⱥ������Ⱥ����
1.CD4���ӵĽṹ��Ϊϸ��Ĥ���浥���ǵ��ף���CD4������458��������л���ɣ������ź���23������л�����Ĥ����374��������л�����2���ǻ����㣬��Ĥ��21������л���������������40������л�����Ĥ��������4��IgV���ṹ���������׳������Ա��
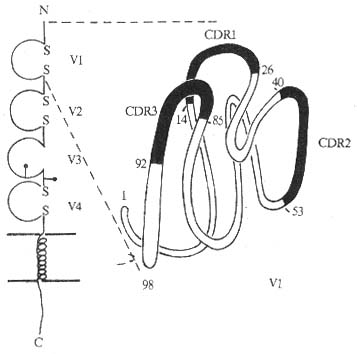
ͼ1-5 CD4���ӽṹģʽͼ
��һ��V������Ig������V���кܸ�ͬԴ�ԣ���3��������������complementarity-determining region, CDR)������3��V������������Poly IgR��ͬԴ����ӽ������е�2��4��V�������������װ���ľ���ֱ�Ϊ28��42��������л�����3��V�����������CD4��Ĥ����MHC������Ӧ����Ŀ�Ĥ���߶�ͬԴ��������CD4����λ�ڵ�2��Ⱦɫ�壬С���6��Ⱦɫ�壬С��CD4���ӵķ�����Ϊ55kDa����457��������л���ɣ��ź�����22��������л���N�˹�����110������Ĥ���һ�������У�long sequence)������262������л�����4���ǻ����㣬��Ĥ��25������л�������������38��������л����˺�С��CD4����Լ��55%������ͬ������������Ϊ�ԡ��ڰ�����λ��3��˿����л���������ΪPKC���ữ�ĵ��CD4�������ֹ������߶ȵı����Ա�����һ����Ĺ�������Ҫ�ġ�
2.CD4���ӵķֲ����ֲ��ڲ���T�ܰ�ϸ��������ϸ�����棬Ҳ������ijЩB�ܰ�ϸ����EBVת����Bϸ������������ϸ������ϸ���������࣬OKT4��Leu3McAb�ɼ��CD4��ԭ��С��L3T4����OKT4����ͬ�
3.CD4���ӵĹ��ܡ��ڳ��������ϸ��������Ѫ����Χ�ܰ������У�CD4����ϸ��һ��Ϊ������T�ܰ�ϸ���յ�ϸ��/������T�ܰ�ϸ���յ�ϸ����helper inducer/suppressor inducer)��
(1)��Ϊϸ����ϸ��֮���ճ�����ӣ�CD4��1��2��������MHC������ӵķǶ�̬���ֽ�����ȶ�MHC����������Ƶ�Tϸ�������MHC��������뿹ԭ�������APCϸ������á���CD4McAb�ɷ��Tϸ���ĸ������ԡ�
��2��ת���źţ�CD4���Ӱ������뵰���Ұ��ἤøp56lck��������Tϸ���źŵ�ת������Ҫ���ã�����ڰ��£���
CD4���Ӱ�Ĥ���1���ṹ������HIV��ǵ���gp120��ʶ��λ������CDR2��gp120��ϵ�������ߣ�CDR3������HIV��Ⱦ��ϸ��Ĥ�ں��йء������� gp120��ϵ�CD4�ķ�Ӧ�ɱ������Լ�����ϣ���1�����CD4V1����CDR2��CDR3��McAb����2��CDR2��CDR3�ĶΣ���3��������CD4 V1�ĶΣ���4����gp120���塣HIV��Ⱦ���������ѡ����CD4+ϸ�����������ٺ��ܽ��ͣ���Ҫͨ�����²�ͬ�Ļ�������1��HIVgp120��Tϸ������CD4���ӽ�Ϻ�ͨ������ѿ���ƻ�ϸ��Ĥ���ڸ�Ⱦϸ�����ڲ������������ϵIJ���RNAֱ������ϸ��Ĥ������ϸ����л��Ӱ��CD4������ϸ��Ĥ�ϵı����Լ��γɶ����ĺϰ��壻��2�����CD4+Tϸ����M��ϸ������MHC���ԭ�Ľ�ϣ�Ӱ��Thϸ���Կ�ԭ��ʶ����̣���3��������������CD4ϸ�������������gp120������HIV�ɷݵĿ��壬ͨ��������ADCCЧӦ����CD4����ϸ������4��������CTLҲ��ͨ��ʶ��CD4ϸ�������gp120���Ӷ�ɱ��CD4����ϸ����������֣�CD26������HIV����һ�����塣
Ӧ�û������������������CD4��rsCD4������ARC��AIDsrelated complexes)�����̲����ڽ��Т����ٴ����飻��CD4McAb��Leu3a)Ҳ�ѿ�ʼ����HIV��Ⱦ��I���ٴ����顣���⣬Ӧ��CD4-IgG��CD4-PE����ŧ�˾��ⶾ�أ���CD4-RA�����鶾��A�����ӽ�����ɱ��HIV��Ⱦ��Tϸ������Ϊ�����̲�����ҩҲ�ѽ����ٴ���֤��1991��������ʪ��ѧ����ϱ������ÿ�CD4McAb�������ʪ�Թؽ��ף�RA���������ƺ��ٴ�֢״���Ը��ƣ�PBMC��CD4����ϸ���ı�����CD4��ԭ�ܶ������½���Ѫ�������CD4��sCD4��ˮƽ�������ߣ�Ѫ����CRP��RF������������ˮƽ���Խ��͡���CD4Ƕ�Ͽ��壨Centocor��˾���������ʪ�Թؽ��ס����Ӳ��֢Ҳ�ѽ�������ٴ���֤����CD4McAb��Ortho Biotech��˾��Ԥ��������ֲ�ųⷴӦ�ѿ�ʼ�ٴ���֤��
���ģ�CD8
1.CD8���ӵĽṹ��CD8�������ɦ�����������������ɵĴ�Ĥ�ǵ��ף�����������34kDa���൱��С���Lyt-2;����30kDa���൱��С���Lyt-3��ÿ����������1��IgV���ṹ�������ġ���Ĥ���Ͱ����������ͦ����������Ĵ��ж��������������CD8��������ͬԴ����˫�壨��/������ɣ�����CD8���Ե�TCR�æ�Tϸ�����档�б�������ϸ���ϵ�CD8����Ϊ�ľ��塣CD8���ͦ���IgV����Լ��110������л�����Ig�ʡ���������V����30��35%ͬԴ�ԣ���V��20��22%ͬԴ�ԣ���TCr V����V����24%ͬԴ�ԡ�����CD8���������Ļ�����Ig�������Ա�������Ig��������������������λ�ڵ�2��Ⱦɫ�壬����ǰ����Ҫ���š�����С��Lyt-2��Lyt -3����λ�ڵ�6��Ⱦɫ�壬���ж�����λ��Ly2a��Ly2b��Ly3a��Ly3b,�ֱ����Lyt-2.1��Lyt-2.2��Lyt-3.1��Lyt-3.2��
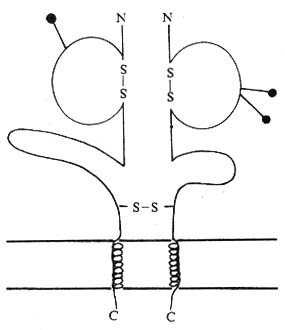
ͼ1-6 CD8���ӽṹģʽͼ
2.CD8���ӵķֲ����ֲ��ڲ���T�ܰ�ϸ��������ϸ����������������ֲ�����пɳ���TCR�æ�CD8��/�����͵�Tϸ����NKϸ�������CD8����Ϊ��/�������塣
������֣�ͨ��ϸ�����ڻ�/�Ͱ�Ĥ���������Ļ��ƣ���Ѫ���д����ſ�����CD8���ӣ�sCD8������Ѫ�����νܽ��ϲ������̲������Դ�Ⱦ�Ե���ϸ������֢�������ϰ���ƶѪ��ͬ��������ֲ�����ʪ�Թؽ���ȫ���Ժ���Ǵ��Ȼ���Ѫ����sCD8ˮƽ���ߣ������ߵ�ˮƽ�뼲�������س̶ȡ�����仯�����Ʒ�Ӧ�Լ�Ԥ���н����еĹ�ϵ��
3.CD8���ӵĹ���
��1����Ϊϸ����ϸ����ĸ�ճ����MHC���ԭ��CD8���ӵ����塣CD8������MHC������ӽ�Ͽ����ȶ�MHC����������Ƶ�Tϸ������Ҫ��CTL�������MHC��������뿹ԭ������İ�ϸ����ϡ�CD8����ϸ��Ϊ������T�ܰ�ϸ��/ɱ����T�ܰ�ϸ����suppressor T lymphocyte/cytotoxic Tlymphocyte,Ts/Tc)��T8��Leu2 McAbʶ��CD8�������ɷ��Tc�Ļ��ԡ�
��2��ת���źţ�Ŀǰ����CD8Ҳ�뵰���Ұ��ἤøp56lck��ء���Tϸ����ֳ�ͷֻ����ź�ת��������Ҫ���á�
��CD8McAb��Ԥ��������ֲʱ��ֲ�������Ӧ������Becton-Dicknson��˾����Leu2�ѽ�������ٴ���֤��
���壩CD2
1.CD2���ӵĽṹ�ͷֲ���CD2�����ֳ�T11�������ϸ�����壨ER�����ܰ�ϸ��������ؿ�ԭ2��LFA-2����Leu5������T�ܰ�ϸ������ĵ����ǵ��ף�������50kDa��CD2����λ�ڵ�1��Ⱦɫ�壬�����������塣����351������л��������ȵ�����24������л���2�����������C2����185������л�����3���ǻ���λ�㣬��Ĥ���Ͱ������ֱַ�Ϊ26��116��������л��������������ΰ���ͼ������ᣬ���������źŵĴ��ݿ����йء���DNAˮƽ���˺�С��CD2��51%ͬԴ�ԡ�CD2���ӷֲ���95%��Tϸ����50��70%����ϸ���ʹ�����ܰ�ϸ����LGL/NK����
2.CD2���ӵĹ���
��1��ճ�����ܣ�CD2���ӵ�������LFA-3�����߷ֲ��ڶ���ϸ�����档CD2����Tϸ���ɽ�Ϻ��д���LFA-3��֬���塣CD2���ӵĹ�����Ҫ��ͨ����CD2McAb���ܰ�ϸ�����ܵ�Ӱ��ͻ���תȾ�������о��ġ��ÿ�CD2McAb�ɣ�1������lectin��ͬ�����忹ԭ�������Կ�ԭ���յ�Tϸ������ֳ��Ӧ����2������T�ܰ�ϸ��IL-2�ϳɺͷ��ڣ���3������CTLЧӦ��ɱ�˹��ܺ�NKϸ����ɱ�˻��ԡ�CTL���ϸ��֮��Ŀ�ԭ��������ճ���ж���;������CD2��LFA-����ϣ�LFA-1��ICAM-1��ϡ��ÿ�CD2�Ϳ�LFA-1���ֿ��壬��LFA-1�Ϳ�LFA-3���ֿ������ȫ���ƿ�ԭ��������ճ�����ã�����CD2�뿹LFA-3���ֿ���ֻ�ܲ�����������ճ�����á�CD2��LFA-3֮���ճ�����ܶ���T�ܰ�ϸ��TCRʶ��������ԭ��APCϸ������MHC��ԭ�����������ԭ��������Ⱦ��ϸ���Լ�ͬ�����忹ԭ������Ҫ�ĸ������á�����о�������CD48��CD59Ҳ��CD2�����壬����Tϸ����ճ����ϸ���������á�
��2��Tϸ����·����;����the alternative pathway of T cellactivation): Reinherz�ɹ����Ʊ���ʶ����CD2������ͬ��λ��McAbs����1��T111��LFA-3��CD58���Ľ���йأ����������ϸ��SRBC����CD58��ͬԴ�T111����SRBC����γ�E��������T111McAb������E�����γɣ���2��T112��CD58����أ���T112McAb���յ�T113��λ���֣���3��T113�ǻTϸ������ı�λ��������ΪCD2R�������յ�T113��λ���ֵĹ����У����뵰���ʺϳ����Ƽ�T113���ܱ������T113���ֲ�����һ���ºϳɵĵ����ʣ��������ڻ���ͱ仯��¶�����ı�λ��ͬʱ���뿹T112��T113McAbs�ɻT�ܰ�ϸ�����ٽ�MHC���ԭ��IL-2����ı������IL-2����ı������IL-2���������£����Tϸ��������ֳ����ΪTϸ����·����;����T113McAb�����IL-3��SRBCͬTϸ��һ�������Ҳ�ܴ̼�Tϸ����ֳ��CD3���Ե�����ϸ����NKϸ������CD2��������⣬CD2����������Ĥ������CD44��CD45�Ĺ����йء�
Ŀǰ����CD2�������ѧ���廹��ʮ�������CD2��������ϸ������LFA-3���Ե�������Ƥϸ����ϣ������������ĵ����£�����Ϊ��������ϸ���Ļ�ṩ�źţ���������ϸ������ֳ�ͷֻ��йء�
������CD58��LFA-3��
1.LFA-3���ӵĽṹ���ܰ�ϸ��������ؿ�ԭ-3��lymphocyte functionassociated antigen-3,LFA-3)��CD58����ϸ�������ǵ��ף�������55��70kDa���������׳������Ա����CD2���Ӹ߶�ͬԴ����Ĥ������2��Ig������C2������CD2��CD58����λ��1��Ⱦɫ�壬�����������������Ǵ�ͬһ�������ƶ�����CD2��CD58�Ľ����������ͬ�ֵ�����ã�homophilic interaction)��
2.LFA-3���ӵķֲ����ֲ���Tϸ����Bϸ��������ϸ������Ƥϸ������Ƥϸ���������֯������άϸ����������ϸ����ѪС����档
3.LFA-3���ӵĹ��ܡ�LFA-3���ӵĹ�����Ҫ��ͨ��Ӧ����Ӧ��McAb����֪����LFA-3����ɣ�1������ЧӦ��CTL�Ļ��ԣ�����������CTL���ϸ��֮���ճ���йأ���2������lectin��ͬ�����忹ԭ�յ���Tϸ����ֳ��Ӧ����EB������Ⱦ��Burkitt���ܰ���ϸ�����У������е�����ϸ������ȱ��LFA-3�ı�����ֿ�CTL��ɱ�����ã���ʾLFA-3ȱ�������ϸ�����������ݻ�����������йء�������ҹ��Ѫ������֢��paroxysmalmocturnal hemoglobinuria,PNH�����ߺ�ϸ������ȱ��LFA-3��DAF��Fc��R��
LFA-3��������ʽ������ϸ��Ĥ���棺��1����Ĥ��ʽ����Ĥ��������Ĥ���Ͱ������ֱ�Ϊ188��23��12���������2��GPI��ê����ʽ����EBVת����JYBϸ��ϵϸ������LFA-3���Ӿ��д�Ĥ��GPI��ê��������ʽ��
[GPI��ê��]�ǻ���֬��������glycosylphosphatidylinsitol,GPI)����Ĥ����ɳɷ֣�GPI�Ǽ��ϵ��Ҵ���ͨ���������̶��ڵ����ʵ��Ȼ��ˣ���Ϊ�����ʶ�λ��ϸ��Ĥ�ϵ�ê�����ֽ�ϵ�GPI�ĵ����ʼ���ΪGPIê���ס�ȥ�ۼ��ܽ�֬��˫��ɻ�ý����GPI�ϵĵ����ʣ���֬øC��PLC���ɴ�ϸ�����潫��һ������֬ê���ĵ������ͷų�������֬��������PLC������������������ἡ����IP3���Ͷ��������ͣ�DG������Ҫ�ĵڶ���ʹ��
���ߣ�CD28
1.CD28���ӵĽṹ��1980��Hara�������õ���¡����9.3������CD28��CD28������44kDa���Ľ�������ɵ�ͬԴ�����壬������Ϊ90kDa���������CD28���ӵ�������202�������ᣬ���а�Ĥ������134�����ᣬ�������׳������Ա����һ��IgV����������С��CD28���ӵ�ͬԴ��Ϊ68%��CD28��CTLA-4�����и߶�ͬԴ��������Ҫ�����ڻ��CTLϸ�����档
2.CD28���ӵķֲ���������Ѫ�ܰ�ϸ����CD28+ϸ��ռ54��86%������90%CD4+Tϸ����50%CD8+Tϸ������CD28��CD28��CD28+Tϸ���б����빦����һ���Ĺ�ϵ��CD8+CD28+Tϸ�����ֳ�MHC���Ƶ�ϸ�������ܣ���CD8+CD28-ϸ�������ƿ�������Լ�ͬ�����忹ԭ���յ���ϸ����ֳЧӦ�����⣬��ϸ���������ֻBϸ��Ҳ�ɱ���CD28��

ͼ1-7 CD28��CD80���������ʾ��ͼ
3.CD28���ӵĹ���
��1��Tϸ����ĸ����źţ���CD28�ɼ�ǿPHA��ConA����CD2McAb����CD3McAb����ֳ�����ЧӦ������IL-2��IL-3��TNF-����IFN-�¡�GM-�á�GM-CSF��ϸ�����Ӳ���������CTLϸ�������⣬��CD28McAb���յ�Tϸ��IL-13mRNA�ı��
��2��CD28����Ȼ������Bϸ�����ԭB7/BB1��CD80����CD28��CD80�Ľ����T-Bϸ���Э������Ҫ��ʽ�����̼�Bϸ�����
������Bϸ��ʶ��ճ������йص�CD����
��һ��BCR������
Bϸ����ԭ���壨B cellreceptor,BCR)���������������ֲ�ͬ�Ķ�������ɣ���ԭ��ϲ�λ�����������������ɵ�Ĥ����Ig�������ṹ�����⣬��BCR�л�����Ig����Ig�����ֶ�����������ڰ�ϸ���ֻ���ԭ����ר�����ۻ��зֱ�����ΪCD79a��CD79b��������Bϸ������mIgM��ص�Ig����Ig�·ֱ�Ϊ47kDa��37kDa�ǵ��ף����������׳������Ա������Ig����Ig�µĻ���ֱ��Ϊmb-1��B29��Ig����Ig�°�Ĥ���������˴�����һ��Ig���ṹ��Ig����Ig�¾�����Ϊ�����Ұ��ἤø�ĵ��������BCR�ź�ת���йأ���ΪmIgM��mIgD������ֻ��3�������ᣨKVK���������ܵ����Ѱ�Ĥ��Ĵ̼��źŴ��ݵ�ϸ���ڡ�Ig����Ig�°�������β����6�����صİ�����л������������ữ��ʽ������в�ͬø�д��ڵ�SH2��src-homology2)�ṹ���ϡ�
������CD19
CD19��һ������Ig�������Ա��������Ϊ95kDa�Ĵ�Ĥ�ǵ��ף��ֲ���Bϸ�����棬����Ӧ�������������в������CD19��Bϸ������źŵ�ת���йأ���1��CD19����¡������յ������ڶ��ֵ���Ѹ�ٷ������ữ����2��CD19�������ɱ�һ��˿���ἤø�����������ữ����3��CD19��������src��ø����Lyn�ȶ��Ľ�ϡ�������һ��Bϸ������źŷŴ��˫�ؿ�ԭ���ģ�ͣ����ģʽ��ΪB�ܰ�ϸ��BCR/Ig����Ig���뿹ԭ��Ϻ�ʹCD19��CD21��ӽ��γɸ����������ԭ��������ԭ��C3dg�ֱ�BCR��CD21����ϣ�������CD19/CD21����������CD19���ܽ�ϵ�src����Lyn����ʹCD19���Ӱ������Ұ��ᷢ�����ữ���й��ź�ת�����̲μ��ڰ��¡��ܰ�ϸ����������ź�ת���ķ��ӻ�������
������CD21
CD21�ֳ�2�Ͳ������壨complementreceptor type 2,CR2)��EB�������壬�Dz��弤����ڼ������һԱ��
[���弤����ڼ�]���弤����ڼ���regulatorsof complement activation,RCA)�������2��Ѫ������H���Ӻ�C4bp�Լ�4��Ĥ����CR1��CR2��DAF��MCP��RCA���ص��ǣ���1������60��70����������ɵĶ�ͬԴ�ظ�˳��short consensus repeat ,SCR);(2)��ϲ����ѽ�Ƭ��C3b��C4b����3������λ��Ⱦɫ��1q32�����������MCP-CR1-CR2-DAF-C4bp��ʽ��������
1.CD21���ӵĽṹ��Ϊ������140kDa�ĵ����ǵ��ף�N����ϸ���⣬��Ĥ����1005�������ᣬ��ˮ��Ĥ��28�������ᣬ�����������ᣬ������34�������ᡣ��Ĥ�������15��SCR��ÿ��SCR����60��70�����ᣬ��һ����4��Cys�ĹǼܽṹ��C1-C4��C2-C3���γ����������������һ��SCR��״�ṹ��SCR1-2��C3dg�����Ŀ�����EBV����йأ���Ĥ������10������12����N-�Ǽ�����������10���������ữΪλ�㡣
2.CD21�ķֲ�����Ҫ�ֲ��ڳ����Bϸ�����ܰ���������ͻ״ϸ��������Tϸ�������⣬���ʡ������Լ�������Ƥϸ��������CD21��ص�145kDa���ӡ�
3.CD21����
��1���ٽ�Bϸ����ֳ������CD21������������Bϸ������ֳ����Tϸ����Tϸ��Դ�Եͷ�����Bϸ���������ӣ�LMW-BCGF�������£��������弤��Bϸ����ֳʱ��CD21��������ǿ�ҵĴٽ����ã��ۺϵ�C3dg��UV-EBV��CD21�����ɼ�ǿTPA�̼���Bϸ����ֳ����LMW-BCGF�����£�CD21�����ɴ̼�Bϸ����ֳ��C3dg�����Ŀ���������LPS�����С��Bϸ��������ֳ����������C3dg�յ�Rajiϸ��������Ѫ�����ֳ�����������ʾ����ѪBϸ����ijЩ�̼���̼���CD21�鵼��ֳ�źŴٽ�Bϸ������ϸ�����ڡ��������CD23��CD21��һ���µ����塣sCD23������ٽ�IgE���ɣ�����BCGF�����ã�CD23��������CD21��ϣ��Ʋ�CD21������������BCGF�����塣
��2��CD21�鵼EBVת��Bϸ����EBV��CD21��CR2����Ϻ���֬øC��ˮ����֬��������������Ca2+�Ͷ���������ˮƽ���ӣ�����Ƶ�����PKC��ת����EBV����B95-8����ȾBϸ�����뷴ʽ����ף�transactivator)EBNA2��LMP����CD21��CD23������Bϸ��������ˮƽ����CD21��CD23��CD23������γ�sCD23���Է��ڵ�BCGF����CD21��ϲ��CD21���ӵ��Ұ��ᵰ��ø�����ϼ���PKC���Ӷ�����Bϸ����ת������ֳ��
��3��CD21���������䣺��ԭ������ʿ�ԭ�ϸ�����C3dgʱ�������ܰ���������ͻ״ϸ������CD21��ϣ����յ����������������Ҫ���á�
��4�����벹��Ļ��CD21���벹�����;���������Լ�C3b�Ĺ̶�������C3bi�ѽ�ΪC3dg������Ϊ˿���ᵰ��ø�����ӵĸ����ӡ�
���ģ�CD80��B7/BB1��
CD80cDNA�ѿ�¡�ɹ��������CD80������262��������ɣ���Ĥ����216�������ᣬ��Ĥ��27�����ᣬ������19�����ᡣB7��BB1�������ֱ�Ϊ44kDa��46kDa����������IJ�������ͬһ���Ķ����ǻ����̶Ȳ�ͬ����ɡ�CD80�������׳������Ա����Bϸ�����ԭ��IFN-�û�ĵ���ϸ��Ҳ����CD80��CD28��CD80�����壬CD28��CD80�Ľ�Ͽ�ͬʱ����Tϸ����Bϸ���Ļ����T��Bϸ��Э���з�����Ҫ���á�
����������Fc������
Ig������������ԭ�ԵIJ����ΪIgM��IgG��IgA��IgD��IgE���࣬����Ig�IJ�ͬ������Ҫ����ṹ�йء�����������ϸ��������в�ͬ��Ig Fc�����壬ͨ��Fc������Ig Fc�Ľ�ϣ�����Ig�鵼���������ܻ������˹��̡�Ŀǰ�Ѽ�����ȷ����CD��ԭ��Fc������Fc��R��Fc��R��Fc��R��
��һ��Fc��R��CD64��CD32��CD16��
1.Fc��R�Ľṹ�ͷֲ���Fc��R�ɷ�ΪFc��R��Fc��R���Fc��R�����࣬���ǵĽṹ�ͷֲ�������ͬ��
��1��Fc��R��CD64����70kDa��Ĥ�ǵ��ף���Ig�������Ա����Ĥ������3��C2�ṹ������Ⱦɫ�嶨λ��1q23��24��ʶ��CD64�Ĵ�����McAb��McAb22��McAb32.2��197��10.1��Fc��R���Ǹ��������壬KdֵΪ10-8��10-9M����Ҫ���˵ĵ���IgG1��IgG3�Լ�С��IgG2a��IgG3��ϡ�����IgG4��ϵ�������ȷ���ͣ���IgG2�����������Fc��R����Ҫ�ֲ��ڵ���ϸ��������ϸ����������ϸ���ȣ�������ˮƽ������ͬ��Fc��R��λ������15000��40000/ÿ������ϸ����>50000/����ϸ����<1000/����������ϸ����IFN-�ÿɴ̼�����ϸ��������ϸ����������ϸ������Fc��R��ˮƽ����5��10����G-CSFҲ�����ִٽ����á�
��2��Fc��R��CD32����40kDa��Ĥ�ǵ��ף�����Ig�������Ա����Ĥ������2��C2�ṹ������Ⱦɫ�嶨λ��1q23��24��ʶ��CD32�Ĵ�����McAb��CIkM5��IV��3��KuFc79��41H16�ȡ�Fc��R���뵥����IgG1��IgG3��IgG4���Ϊ��������Kd5��10-7M��Fc��R������ڳ���ϸ���������Ѫϸ����������Ŀ��20000��40000/ÿ��ϸ��������DNA���к��ܲ�ͬ��Fc��R��ɷ�Ϊ������ʽ����ͬ��ʽFc��R��IJ����Ҫ���ڰ������Ľṹ��ͬ��
��3��Fc��R��CD16����50��70kDa�ǵ��ף���Ig�������Ա����2��C2�ṹ������Ⱦɫ��λ��1q23��24��ʶ��CD16�����Ե�McAb��HUNK2��Leu11��3G8��Gran1��B73.1�ȡ�Fc��R������IgG��IgG3��Ϊ���������塣Fc��R����Fc��R��A��Fc��R��B�������ͣ���Fc��R��A����Ĥ�ṹ����Ҫ�ֲ��ھ���ϸ����NKϸ������������ϸ��������ϸ�������ˮƽFc��R��A��������ϸ������ˮƽ�ϵ͡�Fc��R��A���������ӵ�CD3�ƻ�Fc��R�����˫����أ�����ϸ����Fc��R��A��CD3�����������أ�NK/LGL��Fc��R��A���������ء�TGF-�´ٽ������ĵ���ϸ������Fc��R��A����Fc��R��B��ͨ��GPI��ê����������ϸ�����棬ÿ����������ϸ������10��20��ѪҺ�п����Ե�Fc��R����Ҫ����������ʽ��������ϸ���������ʱ�䴦��������Խ���Fc��R��B�ı���ˮƽ��������ͨ��������Դ�Ե���ø�г�GPI���ӷ����йء�
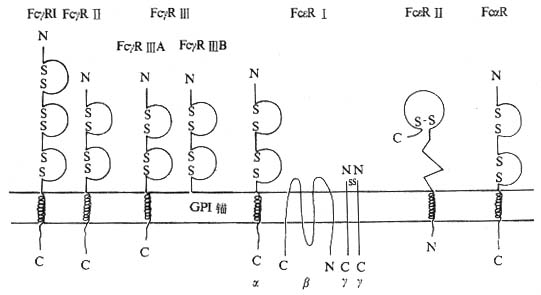
ͼ1-8 Fc��R��Fc��R��Fc��R�ṹʾ��ͼ
2.Fc��R�Ĺ��ܡ�Fc��R�Ĺ�����Ҫ��ͨ������ϸ����NKϸ�������ӵġ�
��1������-����ϸ����Fc��R��͢���ɽ鵼�˵���ϸ��ADCC��ɱ�������Ȱ�ϸ��������ADCCЧӦΪMg2+����������LFA-1��ճ�����Ӳ��롣IFN-�ÿɴٽ�����ϸ��Fc��R��鵼��ɱ�����á�����-����ϸ����ͨ��Fc��R�ӵ������ɺ�������߸���������á�
��2��������ϸ�������ʷ����������ϸ������ͨ��Fc��R�ܽ��ϸ��������IFN-�ô̼��¿�ͨ��Fc��R���Fc��R��鵼ɱ�����ã�����Fc��R��IFN-����Ҫ���յ������ˮƽ���ߣ�����Fc��R�����ˮƽ��δ���ı䣬������ͨ����ɱ�˻����ĵ��ڡ�GM-CSFҲ��ͨ��Fc��R����ȷ����������ϸ����ɱ��ˮƽ��GPI���ӵ�Fc��R��B�����ܽ鵼������ϸ��ɱ�����������á��������ϸ��ͨ��Fc��R�ӵ������ɺ�������߸���������á�
��3����������ϸ����δ�̼�����������ϸ��û��ɱ�����ã�GM-CSF��TNF��IL-5������������ϸ������ADCCЧӦ����Ч���������ɱ�˼�����Ϳ�����������Ҫ���á�GM-CSF����������ϸ���ļ���������Ҫ��ͨ��Fc��R��鵼�ġ�
��4��NKϸ����ͨ��Fc��R��A�鵼ADCCɱ������ϸ���Ȱ�ϸ����IL-2��IFN-�ÿ��������NKϸ����ɱ�˻��ԣ����������Ըı�Fc��R��A�ı���ˮƽ��
������Fc��R��CD89��
Fc��R��CD89��Ϊ������60kDa��Ĥ�ǵ��ף���Ĥ����206�����ᣬ��Ĥ��19�����ᣬ������Ϊ41�����ᣬ��Ig�������Ա����Ĥ����2��C2�ṹ��Ϊ���������壬��Ҫ�����ڵ���ϸ��������ϸ����������ϸ���ȣ��鵼���ɡ�ADCC�Լ���֢���ʵ��ͷš�������ϸ������Fc��R�ɽ��Ѫ���ͺͷ�����IgA1��IgA2������ԼΪKd5��10-7M���Ȼ�ѧ����������IgA�ɴ̼�������ϸ���ѿ�����
������Fc��R��Fc��R��Fc��R��
1.Fc��R�Ľṹ�ͷֲ� Fc��R�ɷ�ΪFc��R���Fc��R�����࣬��ṹ���ֲ����鵼������ѧ����������ͬ��
��1��Fc��R��Ϊ���������壬Kd10-9��10-10M���ɦ����¡���-����������ɡ����Ц�����222��������л���������Ϊ25kDa����Ĥ������Ig������ṹ��2��C2������Fc��R���Fc��R��߶�ͬԴ����Ĥ������IgEC��2/C��3��ϣ���Ĥ��20���Ұ������к�����Fc��R��A��ͬ��8��������л�����������31��������ṹ��Ϊ���ء�������243��������л���������Ϊ33kDa�����ĸ���Ĥ���֣�N����C�˶�λ�ڰ����ڣ��������ܰѦ����ͦ�-��������һ�����������ɶ�����������ͬԴ�����壬ÿ����62�������ᣬ������Ϊ8kDa����Ĥ����ֻ��5��������л���������CD35�߶�ͬԴ��������Fc��R�������ȶ��Ժ��źŵ�ת���йء�NKϸ������Fc��R��A��CD16��������CD3�ƻ�Fc��R�������������ʾFc��R�������CD3�������ЦƵĽṹ���ܵ������ԡ�Fc��R����Ҫ�ֲ����ȼ�����ϸ���ͷʴ�ϸ����
��2��Fc��R��CD23�������������壬������45kDa��������Ĥ�ǵ��ף����Ϳ�Ĥ���ף���C��ֲ��Ѫ���ؼ����Ա��CD23����321�������ᣬN���ڰ�Ĥ�ڣ�1��23λ��������ɰ���β��24��43λ������Ϊ��ˮ��Ĥ������C�˰�Ĥ������277����������ɣ���һ���ǻ����㣬82��102��125��150λ������л�Ϊ����ˮ��ø����λ�㣬������ͬԴ��λ��C��163Cys��282Cys֮�䣬��ͬԴ������6��Cys��88��116λ������֮����һ�������������ṹ������CD23����ͬԴ��������γɡ�CD23���ӿ���Ĥ��C���ѽ�IJ�ͬƬ��14kDa��25kDa��33��37kDaƬ�ξ���ΪIgE������ӣ�IgE-binding factor IgE-BF����Fc��R����ڵ���ˮ��ø�ѽ���γɿ�����CD23���ӣ�sCD23����IgE-BF��CD23mRNA��Fc��R��amRNA��Fc��R��bmRNA���֣������������CD23���ӽ��ڰ�������7��������л��IJ��ͼ1-9����Fc��R��a��Bϸ�����������ΪsCD23��Fc��R��b������Bϸ����Tϸ������������ϸ����ѪС�塢����ϸ��������ϸ������ͻ״ϸ�����ʺ���ϸ��������EBV������ı��ʰ�ϸ��������ϸ��ϵ��U937�ȣ���Ҫ��Ĥ������ʽ���ڣ�IgEC��3��Fc��R�����йء�IL-4���յ�����Bϸ��������ϸ������������ϸ��ת¼Fc��R��bmRNA���ٽ�CD23�ĺϳ�����EBV�˵���EBNA2��CD23���PsCD23���ͷ�Ҳ�дٽ����á���IFN-�á�TGF-�¡�PGE2����Ƥ�ʼ��صȶ�Bϸ������CD23���ͷ�sCD23�����������á�
2.Fc��R����
��1��Fc��R���ȼ�����ϸ���ͷʴ�ϸ�����и�����Fc��R��ÿ��ϸ������Լ��10���������Ӧ��Ӧԭ�놨������ϸ�����ʴ�ϸ������IgE/Fc��R�����Ϻ�ͨ������ʹ���ἡ��ˮ�⣬����Ca2+Ũ�����ӣ�ʹϸ���ѿ������ϳɺ��ͷ���֯����LT��PAF�ȶ��ֽ��ʣ��鵼�����ٷ��ͳ�����Ӧ��
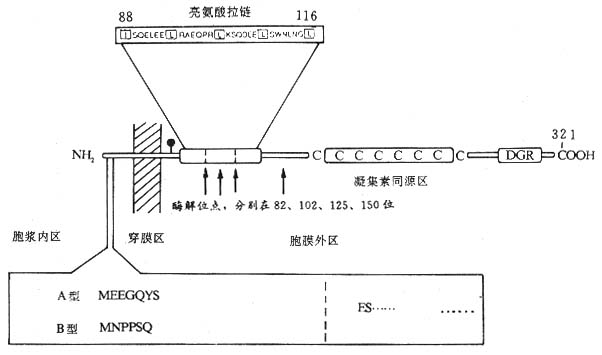
ͼ1-9 CD23���ӽṹģʽͼ
��2��Fc��R��Fc��R��ΪBϸ���������ԭ����̬��Ӧ�Լ�������PBMC��CD23�ܶ��������ӣ�Ѫ��IgE-BP(sCD23�����ߡ�sCD23����Bϸ���������ӣ�bcell growth. factor, BCGF)���ԣ����ֳ�Bϸ����Դ��Bϸ���������ӣ�B-BCGF����sCD23�����������������ÿ�������Ϊ����������CD21��CR2����Ϻ�鵼�ģ�CD23����ͨ���ɽ��̼ˮ�������������ͬԴ����CD21������ϡ����⣬sCD23ͨ�������������ṹ������Bϸ��ĤCD21���ӽ������ٽ�Bϸ��������sCD23��ĤCD23�����������ã��ٽ�Bϸ���ķֻ���IgE�IJ���������IL-4��Эͬ���ã����⣬Fc��R�ɽ鵼IgE������ADCC���������á�CD23��B�ܰ�ϸ����ת��������йأ�EBVת��Bϸ����ֻ��Bϸ������CD23���ܽ�����������ϸ��ϵ��������EBV�˵���EBNA2�йأ�EBNA2�����Fc��R��a������ʼ��λ-275��-89��һ��189bp DNAƬ�ν�ϣ���ʽ������Fc��R��a���������ӣ����յ�Bϸ���߱���CD23��sCD23����ĤCD21���ӽ������γ�һ���Է����������ơ�����B�ܰ�ϸ����Ѫ����B-CLL������Bϸ������CD23���ӣ�����Ѫ����sCD23ˮƽ�������ߡ�
����IgM���塢IgD����Ҳ�б�����ǰ����Ҫ������Bϸ�������߱����ڳ���Bϸ����������Ig Fc������δ����CD�ı�ţ����ڱ��ڴ��ԡ���������IgA��IgM��Ĥת�����������Һ���йصĶ��Ig���壨Poly-IgR),ͨ������Ig�е�J����϶��鵼Igת�˹��ܣ�PolyIgR�������׳������Ա��
����Ȫ��