第一节 独特型和抗独特型
一、独特型发现
本世纪中叶,人们发现将Ig注入同种或异种动物体内可诱导产生抗Ig的抗血清。随后发现了在Ig分子上有两类抗原决定簇,即同种型(isotype)和同种异型(allotype)。1955年Slater等在研究兔抗人骨髓瘤蛋白抗血清的特异性时,发现抗某一些骨髓瘤蛋白抗血清用无Slater等在研究兔抗人骨髓瘤蛋白抗血清的特异性时,发现抗某一些骨髓瘤蛋白抗血清用无关的其它骨髓瘤蛋白或正常人Ig充分吸收后,仍然与最初用作免疫原的那种骨髓瘤蛋白起反应,他们把存在于Ig上这种新的抗原特异性称为“个体”抗原特异性(‘individual'antigenicspecitficities)。
1963年Oudin等用伤寒沙门氏菌免疫50只家兔,将其中一只家兔血清中分离的抗伤寒沙门氏菌抗体作为免疫原,免疫正常家兔,并分离其血清得到抗抗体。研究发现,这种抗抗体只能与作为免疫原的抗伤寒少门氏菌抗体发生常常反应,但不能与其余49只伤寒沙门氏菌免疫家兔的血清发生沉淀反应,也不能与其它抗原免疫家兔血清或正常家兔血清发生反应。上述结果表明,这只家兔产生的抗伤寒沙门氏菌抗体具有特殊的抗原决定簇,它既不同于其它个体(家兔)针对同一抗原(伤寒沙门氏菌)所产生抗体分子上所具有的抗原决定簇,也不同于同一个体针对不同抗原所产生抗体分子上的抗原决定簇,Oudin将这种不同于同种型(isotype)和同种异型(allotype)的抗原决定簇称为idiotype,意为个体基因型。后来人们发现,在在多数情况下,不同个体针对同一抗原的抗体存在着交叉反应,因此不再认为idiotype有严格的个体差异。idiotype现译为独特型。随着名次化学的研究进展,进一步了解到独特型存在于Fab段,它是编码抗体可变区基因的标记。
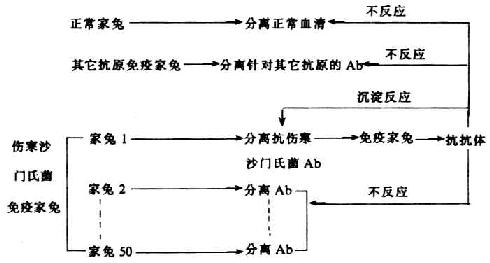
图9-1 独特型抗原特异实验(Oudin,1963)
二、独特的分类
独特型主要是通过抗独特型抗体(anti-idiotypicantibodies,αId)的血清方法来进行分类的。
(一)结合点相关独特型和骨架区相关独特型
通过观察用αId对抗体与相应抗原分子结合是否有抑制作用,可将独特型分为两类:对抗原和抗体的结合有抑制作用的独特型,称为结合点相关独特型(idiotypes associated with combining site);对抗原和抗体的结合无抑制作用的独特型,称为骨架区相关的独特型(framework associated idiotypes)。
(二)私有独特型和交叉反应性独特型
1.私有(或个体)独特型 私有独特型(individual Id,IdI)是指某一特异的抗体分子所特有的独特型,一般认为V区中D基因片段所编码的CDR3与此关系较为密切。
2.交叉反应性独特型 交叉反应性独特型(cross-reactive idiotype,CRI或IdX)是指来自不同个体但具有相同特异性Ab或TCR上的独特型,也可以是来自不同种动物具有相同抗原特异性抗体上的独特型。V基因片段编码的CDR2区域中的某些氨基酸序列(如54、55位氨基酸)常与IdX有关,极少数IdX与重链骨架区和J基因片段所编码的氨基酸序列有关。例如:用葡聚糖(Dextran)免疫BALB/c小鼠制备的两组抗葡聚糖单克隆抗体J558和M104,所有单克隆抗体轻链为λ型,这两组单克隆抗体之间除具有IdX外,各自还具有IdI。它们的交叉反应独特型定位于可变区中的HR(CDR)2;而IdI则定位于HR(CDR)3。换言之,两者在HR2具有共同的抗原决定簇,而在HR3的抗原性彼此不同(图9-2)。
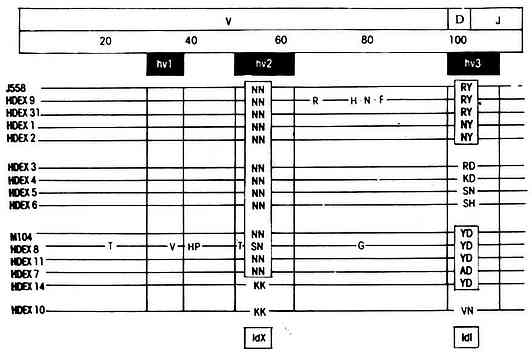
图9-2 抗葡聚糖抗体上的IdX和IdI与结构的相关性
注: N:天冬酰氨酸 Y:酪氨酸 H:组氨酸 M:甲硫氨酸 S:丝氨酸
D:天冬氨酸 R:精氨酸 K:赖氨酸V:缬氨酸
根据IdX与表达IdX动物种系关系的不同,Bona等又反CRI(IdX)分为以下三类。
(1)与同种异型相关的IdX:这种交叉反应独特型表达于所有近交品系或重组近交品系动物相同的同种异型Ig重链或轻链上。
(2)品系间独特型(interstrain idiotypes):这种交叉反应独特型存在于多种近交品系或者一种远系繁殖种中大部分个体。
(3)种间独特型(interspecies idiotypes):不同种不同个体针对相同抗原特异性抗体上的IdX,种间独特型是生物进化过程中胚系基因保守的标记。例如小鼠和大鼠对人工合成抗原GAT的抗体有交叉反应性独特型。再如小鼠、兔和人针对肾综合征出血热病毒抗体上也存在这种类型的IdX。
(三)常规性独特型和调节性独特型
1.常规性(或传统性)Id(conventional idiotypes) 这种Id在机体免疫应答过程中不发生变化,主要存在于天然抗体和自身抗原受体上,前者是由遗传获得,后者在个体发生时已产生了免疫耐受。因此这种Id在机体内的含量很低,不能激活它们的互补性克隆的增殖,在免疫调节上也无重要作用。
2.调节性Id(regulatory idiotypes) 这种Id主要存在于外源性抗原刺激产生的抗体分子上,可激活互补克隆的增殖,在机体免疫应答过程中可发生改变,并参与机体对抗原应答的调节作用。
三 、独特型的分布
1974年Jerne提出把抗原决定簇称为表位(epitope,E),抗体分子上与E互补的结合部位称为补位(paratope,P)。Jerne假设一个表位可被若干种补位以不同的精密度所识别,同样,一种补位可被几种不同的表位所结合。Jerne把抗体分子上V区内存在的若干个表位称为独特位(idiotope,i)如il、i2……等,并认为抗体的独特型是由抗体分子上所有独特位所组成。抗体分子上每个Fab段都有一个补位和一组数量约5-6个的独特位(图9-3)。

图9-3 抗体分子上独特位、独特型、补位以及与抗原表位相互作用的示意图
注:(a)抗体的补位与抗原表位相结合,独特位a、b、c、d、e和f组成独特型
(b)一种补位可结合几种不同表位
(c) 一种补位可结合几种不同补位
应用免疫印迹方法,或免疫化学方法所制备的重链-轻链杂交分子,以及免疫球蛋白可变区蛋白序列的合成肽研究表明,Ig分子特定的独特位主要以单独存在于重链或者单独存在于轻链的方式存在。这在天然Ig分子中也能见到,如E-109骨髓瘤蛋白的Id主要由轻链决定,而A4骨髓瘤蛋白Id主要由重链组成。但在有的情况下,Id分子上的Id依赖于重链和轻链共同组成的天然构象。
四、抗独特型的分类
Bona等根据Id与抗Id的血清学反应和Ab2的功能,将Ab2分为四种类型(图9-4)。由于这种分类方法的实验依据充分,与实际应用联系密切,故应用较为广泛。
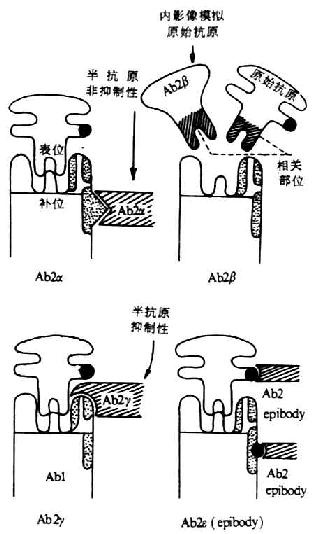
图9-4 Bona抗Id分类模式图
1.Ab2α 这类抗体能识别Ab上与免疫球蛋白骨架区(framework)相关的独特型决定位,Ab2α与Ab1的结合不影响抗原与Ab1的结合,属半抗原非抑制性Ab2(hepten-noninhibitable Ab2)。Ab2α具有调节作用,促进或抑制Ab1克隆。例如抗流感病毒神经氨酸酶特异性抗体所共有的IdX的单克隆抗体,具有促进动物特异性免疫应答,但Ab2α本身并不能诱导动物产生抗神经氨酸酶抗体。
2.Ab2β 具有抗原“内影像”(internal image)的作用,可模拟抗原诱导机体产生针对始动抗原的特异性抗体或细胞免疫应答,因而可抑制Ab1与相应抗原的结合反应,Ab2β在体外可与Ab1上IdX相结合。
已有实验表明,某些Ab2β与模拟的抗原之间的特定部位有较高的同源性,Abβ2可诱导出特异性与Ab1相同的Ab3,这为在分子水平上Abβ2模拟原始抗原提供了证据。需要注意的是,除了蛋白质一级结构外,分子三维结构的相似性,或者与Ab1补位结合点的相似性也可能是Ab2β模拟抗原的分子基础。例如某些b2β可模拟非蛋白质的抗原分子,如细菌的果聚糖、磷酸胆碱、醛固酮、糖脂性质的肿瘤相关抗原等,这可能是b2β某个区域原子核和电子的排列与抗原表位相似,因此可以相似的方式与Ab1的补位相结合。
3.Ab2γ 这类Ab可识别Ab1上与补位(paratope)相关的独特位,能抑制抗原与Ab1的结合,属于半抗原抑制性Ab2(hapten-inhibitable Ab2)。Ab2γ也具有调节作用,可促进或抑制带有相应Id克隆细胞增殖。
4.Ab2ε 又称epibody, 一种双特异性抗体,它能识别Ab1骨架区上的抗原决定簇,同时识别自身或外来抗原上的抗原表位。这种抗独特型抗体对于自身免疫性疾病的研究可能具有意义。