第三节 载脂蛋白B基因多态性的检测
ApoB是富含胆固醇和甘油三酯脂蛋白的重要组成成分,对脂质在人体转运、代谢、维持体内脂质水平的恒定起重要作用。大量临床资料及流行病学调查表明:血清ApoB及低密度脂蛋白(LDL)水平与动脉粥样硬化呈正相关,是其主要危险因素之一。近十年来,从分子水平研究ApoB,阐明动脉粥样硬化及高脂血症的发病机理,寻找其遗传标志受到广泛重视,现已发现:ApoB基因具有明显的多态性,其核苷酸变异有75处,其中导致氨基酸变异的有54处,例如多种酶限制性片段长度多态性(RFLPs),3’端高变区的数目可变的串联重复顺序(VNTR),信号肽(SP)多态性。大量研究表明,ApoB的基因多态性与动脉粥样硬化有不同程度的关系。
一、分析ApoB基因多态性的方法
对ApoB基因多态性进行检测、分析的方法很多,如:限制性内切酶的酶谱分析法,PCR直接分析法,PCR产物RFLP分析法,等位基因特异性寡核苷酸(ASO)杂交法等等,其中最多用且简单的方法有PCR产物直接分析法和PCR产物RFLP分析法。
PCR产物直接分析法,是采用合适的引物,通过多次扩增,获得靶基因序列扩增片段,直接分析法片段的大小,可用于分析基因的缺失与插入。这种方法简便、快速、灵敏,可分析ApoB基因VNTR、SP多态性。
基因突变往往会造成限制性内切酶识别位点的改变,如位点丢失或产生新位点,通过PCR扩增某一特定片段,再用限制性内切酶酶切扩增产物,电泳观察片段的大小,这种方法用于检测酶切位点改变的基因突变,可直接判断基因型,是研究ApoB基因PFLPs的较好方法,具有灵敏、简便、可同时对大量样品进行分析等优点。
二、ApoB基因多态性的检测
ApoB基因多态性研究较多的有RFLP、VNTR、SP多态性,分别叙述如下:
1.RFLP的检测
ApoB有10种RFLP,一般采用PCR-RPLPs方法进行分析,研究较多的有第26外显子XbaⅠ、MspⅠ位点,第29外显子EcoRⅠ位点,在Apob DNA上的位置如图19-6,图19-7所示:
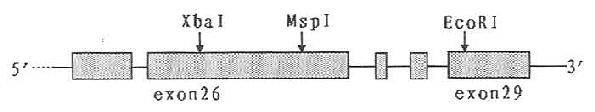
图19-6 ApoB基因部分RFLPS示意图
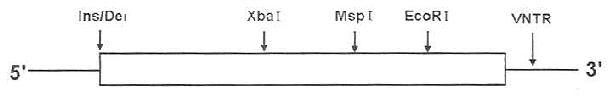
图19-6 ApoB基因多态性位置示意图
用于PCR的引物如下:
XbaⅠ-5’GGAGACTATTCAGAAGCTAA
XbaⅠ-3’GAAGAGCCTGAAGACTGACT
EcoRⅠ-5’CTGAGAGAAGTGTCTTCGAAG
EcoRⅠ-3’CTCGAAAGGAAGTGTAATCAC
MspⅠ-5’TCTCGGGAATATTCAGGAACTATTG
MspⅠ-3’CTAAGGATCCTACAATGTCAAGGT
XbaⅠ酶切位点的RFLP是由于2488位密码子第三个碱基突变(ACC→ACT),产生一个XbaⅠ酶切位点,但并未改变所编码的氨基酸序列,存在XbaⅠ酶切位点(X+)的等位基因,其PCR产物酶切后出现433bp和277bp两个片段,不存在XbaⅠ酶切位点(X-)的等位基因,PCR产物经酶切后仍为其产物710bp片段。
RFLP是由于4154位密码子改变(GAA→AAA),使原有的EcoRⅠ酶切位点消失,氨基酸序列中赖氨酸取代谷氨酸,存在EcoRⅠ酶切位点(R+)的等位基因,其PCR产物经EcoRⅠ酶切后得到253bp及227bp两个片段,不存在EcoRⅠ酶切位点(R-)的等位基因,其PCR产物酶切后仍为480bp的片段。
MspⅠ酶切位点的RFLP,是由于3611位密码子改变(CGG→CAG),谷氨酰胺取代了精氨酸,含MspⅠ酶切位点(M+)的等位基因,PCR产物经MspⅠ酶切后可见到395bp和85bp的片段,不含MspⅠ酶切位点(M-)的等位基因,PCR产物酶切后仍为480bp。
众多学者对ApoB基因RFLPs与动脉粥样硬化之间的关系进行了研究。多数学者认为ApoE有XbaⅠ酶切位点(X+),无EcoRⅠ酶切位点(E-),无MspⅠ酶切位点(M-)的基因频率,在动脉粥样硬化性疾病的患者中比正常对照组高,但也有学者持不同甚至相反意见,究其原因,可能有以下几点:①研究样品例数不够,有的少于100个甚至少于50个,可能使一些有意义的联系被忽略;②对照组的选择没有统一的、严格的、明确的标准,有时与病员组缺乏可比性;③存在种族差异,例如XbaⅠRFLP基因频率,X+等位基因在高加索人为0.4~0.5,日本人和中国人则仅为0.04、0.01;④某些遗传标记之间可能存在连锁关系,可导致结论互不相同。
2.VNTR的检测
VNTR(数目可变的串联重复顺序)是继RFLPs之后的又一类具有高度多态性的遗传标记,广泛分布于人类基因组中。ApoB基因3’端高变区(HVR)包含VNTR,由富含AT的DNA序列组成,其多态性特点是由10~15bp的核苷酸序列构成不同的等位基因,产生不同的重复序列拷贝数。以VNTR两侧特异性序列为引物,经PCR扩增后直接电泳分析产物,片段之间的差异在于AT重复次数不同,重复37次称为3β'37。依此类推,PCR引物可用以下序列:
VNTR-5’ATGGAAACGGAGAAATTATG
VNTR-3’CCTTCTCACTTGGCAAATAC
Boerwinkle等首先检测出12种等位基因片段,现有学者已检出22种不同等位基因。Frield等的研究发现,产生38、44、46或48个15bp长度重复序列的等位基因与冠心病、血浆胆固醇水平升高有关,且与EcoRi RFLP呈连锁不平衡,与MspⅠRFLP呈连锁平衡,Robinson等的研究则没有发现ApoB的VNTR多态性与血浆胆固醇、甘油三酯、ApoB等水平有关。
3.信号肽(SP)多态性的检测
ApoB基因5’信号肽(SP)由27个(插入型Ins)或24个(缺失型Del)氨基酸组成,可经PCR扩增后直接分析,用于PCR的引物序列为:
IN/DE-5’CAGCTGGCGATGGACCCGCCGA
IN/DE-3’ACCGGCCCTGGCGCCCGCCAGCA
Wu等报告del/del基因型在高胆固醇血症患者中比对照组中多见。Renges等对伦敦地区亚洲后裔的研究也发现带del基因型者血浆总胆固醇升高,但与冠心病无明显关系。Peacock等提出Ins/del多态性与冠心病的发展,即其严重程度有关,也有报道Ins/del多态性在冠心病与对照组中无明显差别。
三、ApoB基因多态性检测方法举例
以ApoB基因第26个外显子MspⅠ RFLP为例,具体介绍检测的方法和步骤,笔者对湖北地区健康成人(120例)进行了检测。
(一)材料
引物(如前所述),TaqDNA聚合酶、dNTP、限制性内切酶MspⅠ、高速离心机、基因扩增仪、恒温水浴箱、电泳系统、紫外监测仪。
(二)方法
1.模板DNA的提取
可采用改良的TritonX-100法提取人白细胞DNA,此法结果稳定,但操作较繁琐,在此采用改进的Na裂解法提取人白细胞DNA,简便、快速、灵敏、经济,步骤如下:
取EDTANa2抗凝血100μl,15000r/min离心5min,弃上层血浆,加无菌双蒸水200μl,摇匀20s,加6mNaI200μl,摇匀20s,再加入氯仿/异戊醇(24/1)400μl,摇匀后15000r/min离心12min,小心吸取上层水相,移入另一离心管,加0.6倍体积的异丙醇,混匀后4℃放置15min,15000r/min离心12min,弃异丙醇,加入70%乙醇洗涤2~3次,室温干燥,加入5μlTE缓冲液(pH=8.0)溶解DNA12~24h,4℃备用。
2.模板DNA的扩增
采用聚合酶链反应(PCR)。反应总体积50μl,MgCl2终浓度为1.5mmol/L的1×Buffer,dNTp 0.2mmol/l,引物各20pmol,模板DNA约1.0μg,混匀,短暂离心,95℃预变性10min,加入Taq酶1.5U,液体石蜡50μl,短暂离心,按94℃ 45sec,60℃ 45sec,72℃1mim 15s循环30次,72℃延伸5min,置4℃贮存。
3.扩增产物的提纯及酶切
取扩增产物20μl,加入醋酸铵至2.5mol/L,与无水乙醇混匀,4℃沉淀15min,12000r/min离心10min,弃上清,70%乙醇漂洗两次,室温干燥后加入含Mspi 10U的缓冲液37℃保温2h,加入EDTA至10mmol/L终止反应。
4.扩增及酶切产物的检测
(1)琼脂糖电泳:制备2%琼脂糖(含0.5μg/ml EB),将扩增产物与标准DNA(PGEM3zf(+)/HaeⅢ)点样,电压4v/cm,电泳,紫外灯下观察。
(2)聚丙烯酰胺凝胶电泳:将扩增及酶切产物加样于8%聚丙烯酰胺凝胶上,电压6V/cm,电泳4h,EB染色,紫外灯下观察。
(3)结果:扩增产物经2%琼脂糖电泳,紫外灯下呈一条强荧光带,与已知DNa Marker相比较,约位于480bp的位置,与设计相符。
扩增产物经MspⅠ酶切后,在8%PAGE上,可见到三种类型:①原始带消失,出现两条新带,长度为395bp和85bp,为含酶切位点的M+M+纯合子;②酶切后除与扩增产物相同位置的原始带外,还有两条长度为395bp和85bp的带,共三条带,为M+M-杂合子;③酶切后仅有一条带,与扩增产物位置相同,为无酶切位点的M-M-纯合子。
以上三种基因型,以含酶切位点的纯合型(M+M+)最多见,120例中有114例,不含酶切位点的M-M-型最少见,仅有1例,杂合型(M+M-)有5例,根据基因频率计算方法,等位基因M+频率为0.97,少见等位基因M-频率为0.03。
若对冠心病患者同时进行检测,计算基因频率,可见到:少见等位基因M-在冠心病组比健康对照组高,有显著性差异,但无论在冠心病组还是健康对照组,各基因型个体间的血脂、脂蛋白、载脂蛋白水平均无显著性差异,这说明Apob MspⅠ多态性与冠心病有关,但与血中脂类水平的关系不明显,这可能是由于环境因素,激素及其他遗传因素对脂质水平的影响较明显,掩盖了Apob MspⅠRFLP对脂质的影响,也可能是这种多态性与ApoB基因上其他位点的多态性有连锁关系。另外,由于少见等位基因M-的频率很低,有必要加大样本例数,进行进一步的研究。