�����ڡ������ʵĽṹ���书��
������Ϊ����߷�������֮һ��������ά�ռ�ṹ�����ִ�и��ӵ�����ѧ���ܡ������ʽṹ�빦��֮��Ĺ�ϵ�dz����С����о��У�һ�㽫�����ʷ��ӵĽṹ��Ϊһ���ṹ��ռ�ṹ���ࡣ
һ�������ʵ�һ���ṹ
�����ʵ�һ���ṹ(primarystructure)���ǵ����ʶ������а�����л�������˳��(sequence)��Ҳ�ǵ�����������Ľṹ�������ɻ������Ŵ����������˳���������ġ����ְ����ᰴ�Ŵ������˳��ͨ���ļ�������������Ϊ�����������ļ��ǵ����ʽṹ�е�������
��������Լһǧ�����ҵ����ʵ�һ���ṹ���о�ȷ�������ȵ��أ��Ⱥ��Ǻ���ø���ȵ���ø�ȡ�
�����ʵ�һ���ṹ�����˵����ʵĶ����������ȸ��ṹ���ɰ��ڵ���Ȼ�����ʸ��������������ѧ���ԣ�����ÿһ�ֵ����ʵ�����ѧ���ԵĽṹ�ص㣬���������������İ��������У�������ɵ����ʵ�20�ְ������������IJ������������ŵ��������ʺͿռ��Ų�������ͬ�������ǰ��ղ�ͬ�����й�ϵ���ʱ���Ϳ��γɶ��ֶ����Ŀռ�ṹ�Ͳ�ͬ����ѧ���Եĵ����ʷ��ӡ�
ͼ1-1 �ȵ��ص�һ���ṹ
���������ʵĿռ�ṹ
�����ʷ��ӵĶ��������dz�������չ�������۵��������������еıȽ��ȶ��Ŀռ�ṹ�������ʵ�����ѧ���Ժ�����������Ҫ�����ڿռ�ṹ����������˽����ⶨ�����ʷ��ӵİ�������ɺ����ǵ�����˳������ȫ�˽⵰���ʷ��ӵ�����ѧ���Ժ��������ʡ�������״������(�����Ѫ���еİ��ס��ס�Ѫ�쵰��ø��)����ά״������(�ǵ��ס���ԭ���ס��������ס���ά����)��ǰ������ˮ�����߲�����ˮ���Զ������������ʲ��ܽ��õ����ʵ�һ���ṹ�İ���������˳�������͡�
�����ʵĿռ�ṹ����ָ�����ʵĶ������������ļ��ṹ��
(һ)�����ʵĶ����ṹ
�����ʵĶ����ṹ(secondarystructure)��ָ������������ԭ�ӵľֲ��ռ��Ų��������漰�������ֵĹ���
1.�ļ�ƽ��(�������ƽ�棬amide plane)��
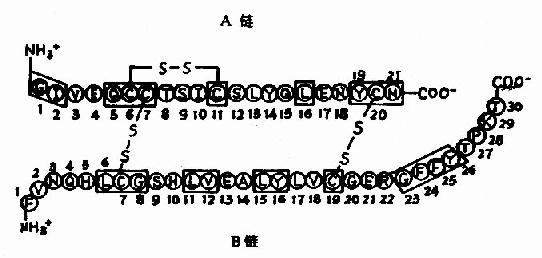
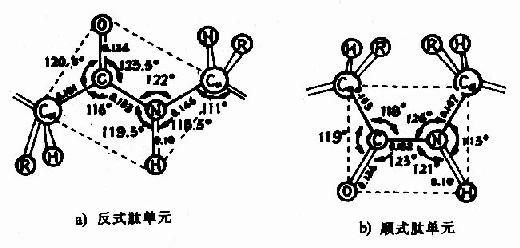
Pauling���˶�һЩ���ļ�������������Ƚ�����X������������ó�ͼ1-2��ʾ�ṹ����һ���ļ�����Χ��������֪��
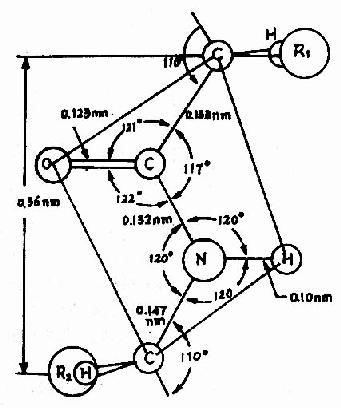
ͼ1-2 �ļ�ƽ��ʾ��ͼ
(1)
(2)
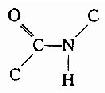
(3)���ļ��е�C-N�Ⱦ���˫�����ʣ��ͻ���˳����ͬ�������칹����֤ʵ
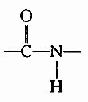
ͼ1-3 ��ʽ�ĵ�Ԫ��˳ʽ�ĵ�Ԫ
2.��������������Ľṹ��Ԫ
1)��������Pauling���˶Ԧ����ǵ���(����keratin)������X�����������������ͼ�п�����0.5��0.55nm���ظ���λ�����Ʋ⵰���ʷ��������ظ��Խṹ������Ϊ�����ظ��ԽṹΪ��������(����helix)��ͼ1-4��
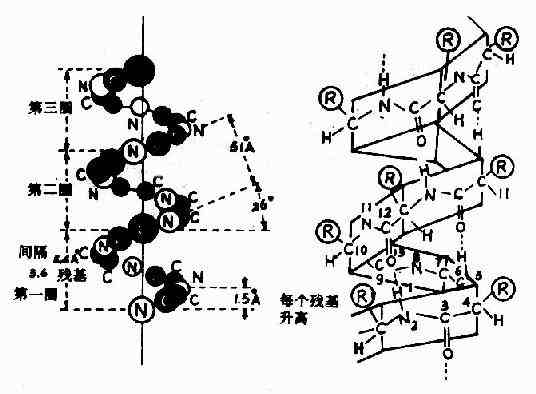
ͼ1-4 �����ʷ��ӵĦ�������
���������Ľṹ�ص����£�
(1)����ļ�ƽ��ͨ������̼ԭ����ת���֮������������ȹ̵�����������
(2)����������������ÿ3.6��������л�����һȦ���൱��0.54nm������X������ͼ���ϡ�
(3)������Ȧ����֮����ļ���C��O��H�N�γ����������⽡����ÿһ��������л��е�NH��ǰ����������л���C��O֮���γ�����������ȶ�������������Ҫ����
(4)�����а��������R���ֲ���������࣬����״����С�����Ӱ������������γɡ����Ի�������Ἧ�е���������ͬ�����⣬�����ڦ��������γɣ��ϴ��R(�籽�����ᡢɫ���ᡢ��������)���е�����Ҳ�������������γɣ��������������̼ԭ��λ����Ԫ���ϣ�����Ťת����֮�����ǰ����ᣬ�����γ�������ʲ����γ����������������ʰ����R��ΪH���ռ�ռλ��С��Ҳ��Ӱ��ô��������ȶ���
2)�£�Ƭ��ṹAstbury�������Ԧ£��ǵ�����X��������������־���0.7nm���ظ���λ���罫ë�������ǵ�����ʪ�����������죬��������ԭ�����������֦���������X������ͼ�ɸı�Ϊ��£��ǵ������Ƶ�����ͼ��˵���£��ǵ����еĽṹ�ͦ�������������չ��ṹ��ͬ���������ϵ������۵��ɾ��״��������ͨ�����������ƽ�г�Ƭ��״�Ľṹ��Ϊ�£�Ƭ��(�£�pleatedsheet)�ṹ��Ʀ£��۵�(ͼ1��5)��
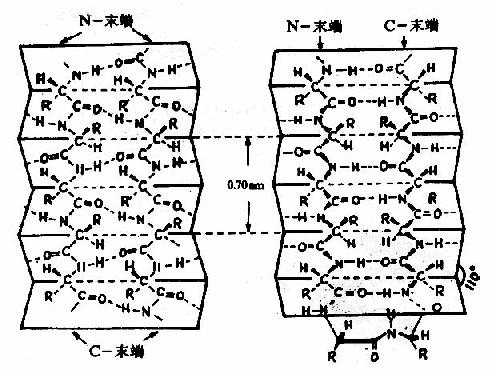
ͼ1-5 �����ʷ����еĦ£�Ƭ��ṹ
��˳��ƽ�С��ң�����ƽ��
�£�Ƭ��ṹ�ص��ǣ�
���������൱��չ�Ľṹ������ƽ��֮���۵��ɾ��״�������ļ�ƽ����110��ǡ�������л���R��������ھ�ݵ��Ϸ����·���
����������������һ�������ڵ������������C��O��H�X�γ������ʹ�����ȶ���
����������������ƽ�еģ�Ҳ�����Ƿ�ƽ�еġ���ǰ���������ӡ�N�ˡ�����C�ˡ���ͬ����ģ������Ƿ�����ġ��£�Ƭ��ṹ����ʽʮ�ֶ�����������ƽ��������档
��ƽ�еĦ£�Ƭ��ṹ�У������л��ļ��Ϊ0.65nm����ƽ�еĦ£�Ƭ��ṹ������Ϊ0.7nm��
3)�£�ת��
�����ʷ����У��������������180��Ļ��ۣ������ֻ��۽Ǵ��Ĺ�����Ǧ£�ת��(�£�turn��£�bend)���£�ת���У���һ��������л���C��O����ĸ��л���N�H�γ�������Ӷ�ʹ�ṹ�ȶ�(ͼ1��6)��
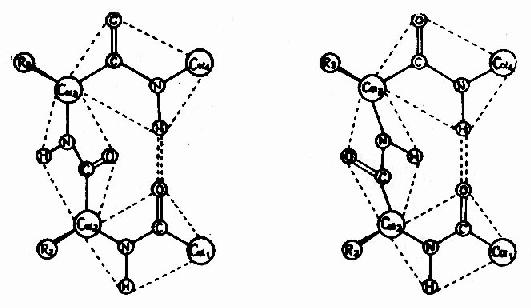
ͼ1-6�������ʷ����еĦ£�ת��
4)�����
û��ȷ�������ԵIJ������������������ļ�ƽ�治�������У�������ɢ�������(random coil)��
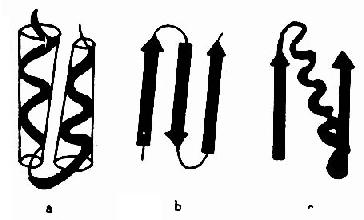
ͼ1-7 �����ʵij������ṹʾ��
a.������ϡ�b.�¦¦���ϡ�c.�¦������
(��)�������ṹ�ͽṹ��
�������ṹ(supersecondarystructure)��ָ�ڶ�������˳������ڽ��Ķ����ṹ�����ڿռ��۵��п������˴�����ã��γɹ���Ķ����ṹ�ۼ��塣Ŀǰ���ֵij������ṹ�����ֻ�����ʽ�����������(����)�����۵����(�¦¦�)�ͦ��������۵����(�¦���)(ͼ1��7)�������Ԧ¦��������Ϊ���������ǿ�ֱ����Ϊ�����ṹ�ġ������顱��ṹ�����ɵ�λ���ǵ����ʹ����ж����ṹ�������ṹ֮���һ����Σ��ʳƳ������ṹ��
�ṹ��(domain)Ҳ�ǵ����ʹ����ж����ṹ�������ṹ֮���һ����Ρ��ڽϴ�ĵ����ʷ����У����ڶ����������ڵij������ṹ������ϵ���γɶ��������ڿռ��Ͽ��������������뵰�����ǻ��ṹ������һ��ÿ���ṹ��Լ��100-200��������л���ɣ����ж��صĿռ乹���е���ͬ������ѧ���ܡ���������(IgG)��12���ṹ����ɣ��������������ϸ���2�������������ϸ���4���������ϲ�λ�뿹ԭ��ϲ�λ���ڲ�ͬ�Ľṹ��һ�������ʷ����еļ����ṹ���е���ͬ���еIJ�ͬ������ͬ�����ʷ���֮�������еĸ��ṹ��Ҳ������ͬ������������ø��3���������ȩ����ø��ƻ��������ø�Ⱦ�����NAD+Ϊ��ø������ø�࣬���Ǹ�����2����ͬ�Ľṹ����ɣ���������NAD+��ϵĽṹ�����������ͬ��
ͼ1-8�������������ṹ��ijЩ�μ���
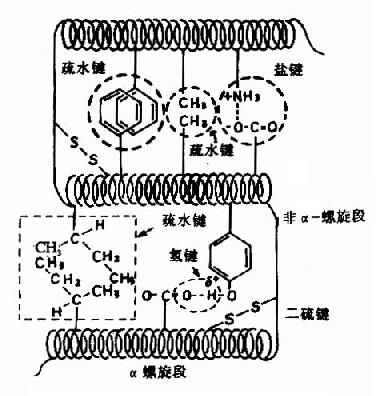
(��)�����ʵ������ṹ
�����ʵĶ������ڸ��ֶ����ṹ�Ļ������ٽ�һ���������۵��γɾ���һ�����ɵ���ά�ռ�ṹ����Ϊ�����ʵ������ṹ(tertiary structure)�������������ṹ���ȶ���Ҫ���μ����������������ˮ�����μ��Լ����»���(Van der Wasls��)��(ͼ1-8)����Щ�μ����ɴ�����һ���ṹ��������Զ�İ�����л���R����֮�䣬��˵����ʵ������ṹ��Ҫָ������л��IJ�����Ľ�ϡ��μ������Ƿǹ��ۼ������ܻ�����pH���¶ȡ�����ǿ�ȵȵ�Ӱ�죬�б䶯�Ŀ����ԡ�����������ڴμ���������ijЩ��������ʹԶ���Ķ����Ķ���ϵ��һ������ڵ����������ṹ���ȶ���������Ҫ���á�
��Ҳ����Ϊ�����ʵ������ṹ��ָ�����ʷ��������۵������γɹ���Ļ����ϣ������еĸ����������γ�һ���Ĺ�����������Ҫ���γ���(��ƽṹ��domain)������״��������˵���γ���ˮ������ˮ������ˮ�����ڵ����ʷ��ӱ��棬�ɺܶ���ˮ������ɡ���ˮ�����ڷ����ڲ�������ˮ�������й��ɣ���ˮ�����γ�һЩ����Ѩ���ڴ�����ijЩ��������Ƕ���У���Ϊ���Բ�λ��
�߱������ṹ�ĵ����ʴ��������Ͽ����е�ϸ��(����ȶ����10������)��������ά״������(fibrous protein)����˿�ĵ��ף��еij�������������ϳ����Σ�������״������(globular protein)����Ѫ���嵰�ס��ס����쵰�ף���״������ˮ����ۼ��ڷ��ӵ��ڲ�������ˮ�����ֲ��ڷ��ӱ��棬�����״����������ˮ�ģ�����Ҫ���ǣ���������������������γ�ijЩ��������ѧ���ܵ��ض���������ø�Ļ������ĵȡ�
(��)�����ʵ��ļ��ṹ
���ж�����������϶��������ṹ�Ķ�������ɵĵ����ʣ����������ͨ���μ������϶��γɵĿռ�ṹ��Ϊ�����ʵ��ļ��ṹ(quarternary structure)�����У�ÿ�����ж��������ṹ�Ķ�������λ��Ϊ�ǻ�(subunit)���ļ��ṹʵ������ָ�ǻ��������Ų�������ü��Ӵ���λ�IJ��֡��ǻ�֮�䲻�����ۼ����ǻ���μ����Ľ�ϱȶ��������ṹ���ɣ������һ���������£��ļ��ṹ�ĵ����ʿɷ���Ϊ����ɵ��ǻ������ǻ����������Կɲ��䡣
һ�ֵ������У��ǻ��ṹ������ͬ��Ҳ�ɲ�ͬ�����̲ݰ��Ʋ�������ǵ�������2200����ͬ���ǻ��γɵĶ���壻������Ѫ�쵰��A���������ǻ����������ǻ��γɵ��ľ��壻�춬���ᰱ������ת��ø�����������ǻ����������ǻ���ɡ����˽�����ȫ�ײ�ͬ�ǻ�����С��λ��Ϊԭ����(protomer)����һ�����ǻ���һ�������ǻ���ϳ��춬���ᰱ������ת��ø��ԭ���塣
ijЩ�����ʷ��ӿɽ�һ���ۺϳɾۺ���(polymer)���ۺ����е��ظ���λ��Ϊ����(monomer)���ۺ���ɰ��������������������ͬ����Ϊ�����塢�����塭���Ѿ���(oligomer)�Ͷ����(polymer)�����ڣ����ȵ���(insulin)�����ڿ��γɶ����弰�����塣
ͼ1��9�����쵰�������ṹ�ͱ��������칹ø�������ṹͼ
ͼ1-10����쵰���ǻ����ģʽͼ
���������ʵĽṹ�빦�ܵĹ�ϵ
(һ)�����ʵ�һ���ṹ���乹���ܵĹ�ϵ
������һ���ṹ�ǿռ�ṹ�Ļ������ض��Ŀռ乹����Ҫ���ɵ����ʷ����������Ͳ���R�����γɵĴμ�����ά�֣����������ڣ������ʵĶ�����һ�����ϳɺ��ɸ���һ���ṹ���ص���Ȼ�۵����������γ�һ���Ŀռ乹��
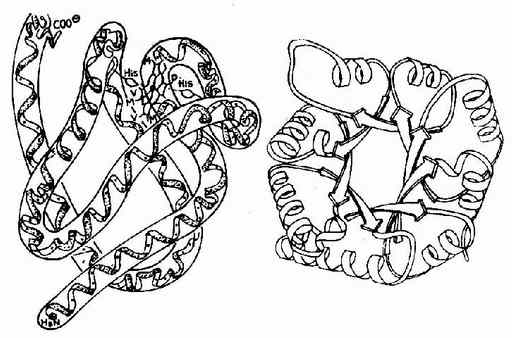
Anfinsen��һ�������ĵ����ʺ��Ǻ���øΪ�����о�������Ļ�ԭ���������⣬���ָ�ø��124��������л����ɵĶ������д����ĶԶ�������ڴ����£��ϻ��Ҵ����������������£��ĶԶ����ȫ������ԭΪ�SH��ø����Ҳȫ��ɥʧ�������罫���غͦ£��ϻ��Ҵ���ȥ����������������ʹ�ϻ����������ɶ��������ʱø�Ļ���ˮƽ�ɽӽ�����Ȼ��ø��Anfinsen�ڴ˻�������Ϊ�����ʵ�һ���ṹ���������Ķ����������ṹ������һ���ṹ�����Զ��ط�չ�����������ṹ(ͼ1-10)��
һ���ṹ���Ƶĵ����ʣ������������Ҳ���ƣ����磬��ͬ��������������������ͬһ���ܵĵ����ʣ���һ���ṹֻ�м��ٵIJ�𣬶�����ϵͳ�����Ͻ���λ����������IJ�����С(��1-2����1��3)��
��1-2���ȵ��ط����а�����л��IJ��첿��
| �ȵ�����Դ | ������л��IJ��첿�� | |||
| A5 | A6 | A10 | A30 | |
| �� | Thr | Ser | Ile | Thr |
| �� | Thr | Ser | Ile | Ala |
| �� | Thr | Ser | Ile | Ala |
| �� | Thr | Ser | Ile | Ser |
| ţ | Ala | Ser | Val | Ala |
| �� | Ala | Gly | Val | Ala |
| �� | Thr | Gly | Ile | Ala |
| Ĩ��S | Thr | Ser | Ile | Ala |
| ��S | Ala | Ser | Thr | Ala |
��1-3��ϸ��ɫ��C�����а�����л��IJ�����Ŀ������ʱ��
| ��ͬ���� | ������л��IJ�����Ŀ | ����ʱ��(������) |
| ��-�� | 1 | 50-60 |
| ��-�� | 12 | 70-75 |
| ��-�� | 10 | 70-75 |
| ��-ţ-�� | 0 | |
| ��-ţ | 3 | 60-65 |
| ������-�� | 10-15 | 280 |
| ������-� | 17-21 | 400 |
| ������-��ĸ | 43-48 | 1,100 |
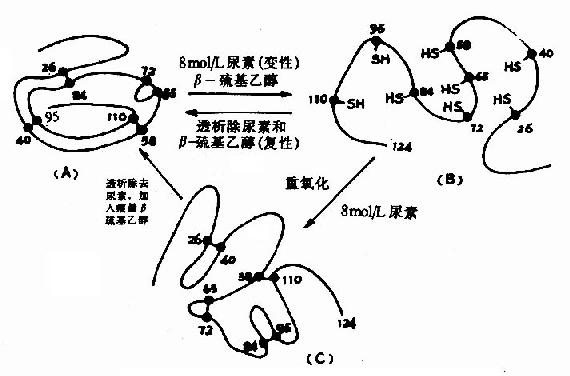
ͼ1-11�����Ǻ���ø�ı��Ժ���ʾ��ͼ
(A)��Ȼ���Ǻ���ø(B)����ʧ��(C)�����ҡ����Ǻ���ø
��������Ƥ�ʼ���(ACTH)�ʹٺڼ���(MSH)��Ϊ������ڵĶ��ļ��ء�����MSH��ACTh 4��10λ�İ�����ṹ��£�MSH��11��17λһ������ACTH�н�����MSH����������(ͼ1-12)��
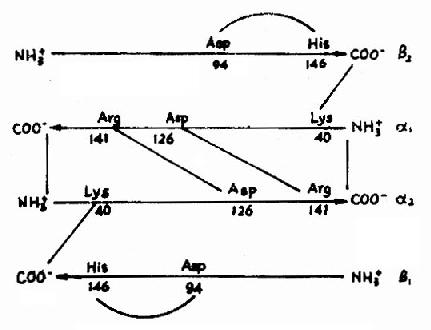
�ڵ����ʵ�һ���ṹ�У����빦�ܻ��Բ�λ�IJл������ض�����ؼ���λ�IJл�����ʹ�����������з���һ���л����쳣����ô�õ����ʵĹ���Ҳ���ܵ����Ե�Ӱ�졣����֮Ϊ�����Ӳ���������״��ϸ����ƶѪ������574��������л��У�һ��������л������ǻ�N�˵ĵ�6�Ű�����л������˱�������ɵģ����ֱ�����Դ�ڻ������Ŵ���Ϣ��ͻ��(��ͼ1-13)��
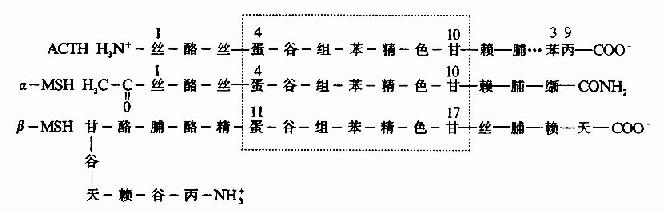
ͼ1-12��ACTH����-MSH�ͦ�-MSHһ���ṹ�Ƚ�
| ���� | DNA | ����TGt GGG CTT CTT TTT���� |
| mRNA | ACA CCC GAA GAA AAA | |
| DNA(���ǻ��� | N�ˡ���-��-��-��-������ | |
| �쳣 | DNA | ����TGT GGG GAT CTT TTT���� |
| mRNA | ����ACa CCC GUA GAA AAA���� | |
| hbs(���ǻ��� | N�ˡ���-��-��-��-������ |
ͼ1-13������״��ϸ����ƶѪѪ�쵰���Ŵ���Ϣ���쳣
(��)�����ʿռ������빦�ܻ��ԵĹ�ϵ
�����ʶ��ֶ����Ĺ�������ֵ������ض��Ŀռ乹��������أ������ʵĿռ乹�����书�ܻ��ԵĻ������������仯���书�ܻ���Ҳ��֮�ı䡣�����ʱ���ʱ��������ռ乹���ƻ����������ܻ���ɥʧ�����Ե������ڸ��Ժ���ԭ�����Լ��ָܻ���
���������ڣ���ij������������뵰���ʷ��ӵ�ij����λ��ϣ������õ����ʵĹ�����һ���仯���Ӷ������书�ܻ��Եı仯�����������Ϊ�����ʵı�ЧӦ(allostery)��
������(��ø)�ı�ЧӦ�������������ձ���ڣ�������ʴ�л�ĵ��ں�ijЩ�������ܵı仯����ʮ����Ҫ�ġ�
����Ѫ�쵰��(hemoglobin,��дHb)Ϊ����˵�������빦�ܵĹ�ϵ��
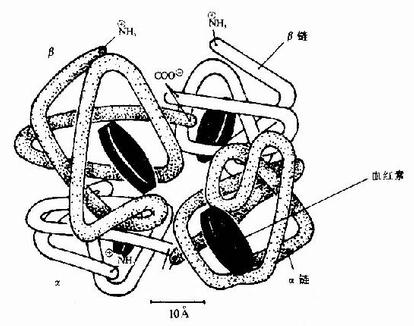
Ѫ�쵰���Ǻ�ϸ���������е�һ�ֽ�ϵ����ʣ����ĵ����ʲ��ֳ�Ϊ�鵰��(globin)���ǵ����ʲ���(����)��ΪѪ����(��ͼ1-14)��Hb�������ĸ��ǻ����ɣ�ÿһ�ǻ����һ����Ѫ���ء���������Hb���ӵ��ĸ��ǻ�Ϊ��������������������������141��������л���ɣ�������146��������л���ɣ����ǵ�һ���ṹ����ȷ����ÿһ�ǻ������ж����������ṹ���������۵�������һ�������ǻ�����8������������(�ֱ��A��B����H������)�����ǻ�����7���������������ڴ˻�����������һ���۵��γ���״�������������γɵĸ��ִμ���ά���ȶ���ʹ֮���α���Ϊ��ˮ�����������ڣ���E��F�����μ��20�����ˮ������������ɿڴ��ε���ˮ��������Ѫ���ؾ�Ƕ�������У����ǻ��ͦ��ǻ��������ƣ�����ĸ��ǻ���2��2�ۺϳɾ����ļ��ṹ��Hb����(��ͼ1-15)���ڴ˷����У��ĸ��ǻ����������Ų��ķ��������ǻ��ز�ͬ����Ƕ���������ǻ��䣬���ǻ��������ִμ�����ϵ��ʹ�������ӳ����Σ���Щ�μ�������άϵHb���ӿռ乹������Ҫ���ã����������ǻ����8���μ�(ͼ1-16)�����ǵ��γɺͶ��ѽ�ʹ�������ӵĿռ乹�����仯��
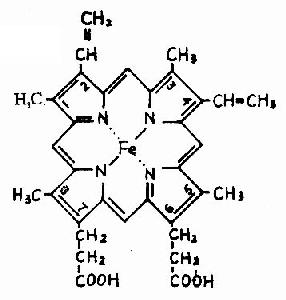
ͼ1-14 Ѫ���صĽṹʽ
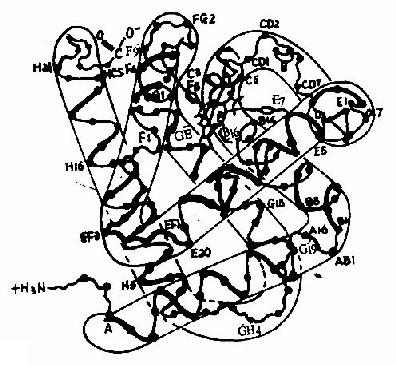
ͼ1-15��Ѫ�쵰�צ��ǻ��Ĺ���
ABCDEFGH�ֱ������ͬ�Ħ��������������а˸������������������ִ����ڸ���������л�����ţ�a-������֮������в�λΪ���������AB��CD��EF��FG���ȱ�ʾ��C1��E7��C5��CF��C3��E3�����м�ΪѪ���أ����нϴ�ĺڵ����Fe2+��
ͼ1-16 Ѫ�쵰���ǻ����μ�ʾ��ͼ
ͼ1-7����ԭ��������ʱ����Ѫ����ƽ��

ͼ1-18
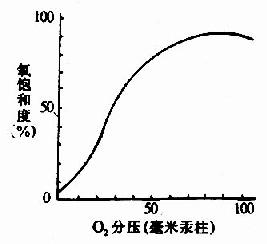
ͼ1-19��Hb������������
Hb�����ڵ���Ҫ����Ϊ������������Hb�ı�λЧӦ�������������ڷβ���O2��ϼ�����Χ��֯�ͷ�O2��
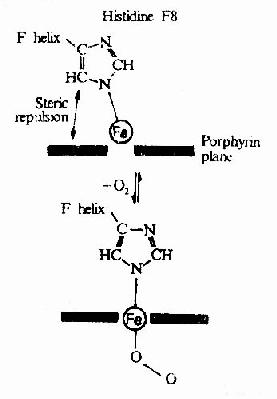
Hb��ͨ���丨��Ѫ���ص�Fe++�������������ϵģ�Ѫ���ص���ԭ�ӹ���6����λ��������4����Ѫ���ص���������N��ϣ�һ�����鵰���ǻ�F�������ĵ�8λ�鰱��(F8)�л����������N�����ӣ����ŵ�һ����λ������O2����ؽ�ϣ�����������Ѫ�쵰�ס�
��Ѫ�����У��ĸ��������γ�һ��ƽ�棬��δ�������ʱFe++��λ�ø���ƽ��0.7Å��һ��O2����ijһ�����ǻ�����ˮ���ڴ���ʱ����Fe++�Ľ�ϻ�ʹFe++Ƕ��������ƽ���У�Ҳ�����ƽ�����ƶ�Լ0.75Å(ͼ1-17)������λ�õ���һС�ƶ���ǣ��F8�鰱��л���ͬF�����ε�λ�ƣ��ٲ��������Ķι�������������ǻ����μ����ѣ�ʹ�ǻ����ϱ��ɣ����ٽ��ڶ��ǻ��ı乹�����ϣ������ִٽ������ǻ�������(ͼ1-18)ʹHb�����е����ǻ��������ٶ�Ϊ��һ�ǻ���ʼ����ʱ�ٶȵ����ٱ�������һ���ǻ��ı����ã��ٽ���һ�ǻ��乹������Ϊ�ǻ����ЭͬЧӦ(cooperativity)�������ڲ�ͬ����ѹ�£�Hb���������߳ʡ�S����(ͼ1-19)��