第三节 嘧啶核苷酸的合成代谢
嘧啶核苷酸合成也有两条途径:即从头合成和补救合成。本节主要论述其从头合成途径。
(一)嘧啶核苷酸的从头合成
与嘌呤合成相比,嘧啶核苷酸的从头合成较简单,同位素示踪证明,构成嘧啶环的N1、C4、C5及C6均由天冬氨酸提供,C3来源于CO2,N3来源于谷氨酰胺。(图8-7)
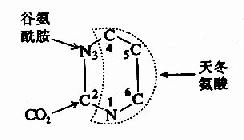
图8-7 嘧啶环合成的原料来源
嘧啶核苷酸的合成是先合成嘧啶环,然后再与磷酸核糖相连而成的。
1.尿嘧啶核苷酸(UMP)的合成,由6步反应完成:(图8-8)
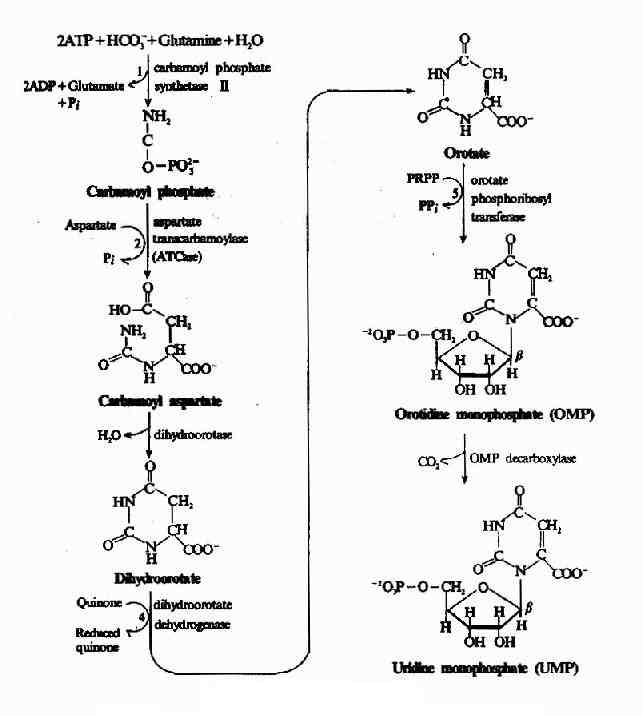
图8-8 UMP的生物合成
(1)合成氨基甲酰磷酸(carbamoyl phosphate):嘧啶合成的第一步是生成氨基甲酰磷酸,由氨基甲酰磷酸合成酶Ⅱ(carbamoyl phosphate synthetase Ⅱ,CPS-Ⅱ)催化CO2与谷氨酰胺的缩合生成。正如氨基酸代谢中所讨论的,氨基甲酰磷酸也是尿素合成的起始原料。但尿素合成中所需氨基甲酰磷酸是在肝线粒体中由CPS-Ⅰ催化合成,以NH3为氮源;而嘧啶合成中的氨基甲酰磷酸在胞液中由CPSⅡ催化生成,利用谷氨酰胺提供氮源。CPS-Ⅰ和CPS-Ⅱ的比较见下表8-1。
(2)合成甲酰天冬氨酸(carbamoyl aspartate):由天冬氨酸氨基甲酰转移酶(aspartate transcarbamoylase,ATCase)催化天冬氨酸与氨基甲酰磷酸缩合,生成氨基甲酰天冬氨酸(carbamoyl aspartate)。此反应为嘧啶合成的限速步骤。ATCase是限速酶,受产物的反馈抑制。不消耗ATP,由氨基甲酰磷酸水解供能。
表8-1 两种氨基甲酰磷酸合成酶的比较
| 氨基甲酰磷酸合成酶Ⅰ | 氨基甲酰磷酸合成酶Ⅱ | |
| 分布 | 线粒体现肝脏) | 胞液(所有细胞) |
| 氮源 | 氨 | 谷氨酰胺 |
| 变构激活剂 | N乙酰谷氨酸 | 无 |
| 反馈抑制剂 | 无 | UMP(哺乳类动物) |
| 功能 | 尿素合成 | 嘧啶合成 |
(3)闭环生成二氢乳清酸(dihydroortate):由二氢乳清酸酶(dihyolroorotase)催化氨基甲酰天冬氨酸脱水、分子内重排形成具有嘧啶环的二氢乳清酸。
(4)二氢乳清酸的氧化:由二氢乳清酸还原酶(dihydroorotate dehyolrogenase)催化,二氢乳清酸氧化生成乳清酸(orotate)。此酶需FMN和非血红素Fe2+,位于线粒体内膜的外侧面,由醌类(quinones)提供氧化能力,嘧啶合成中的其余5种酶均存在于胞液中。
(5)获得磷酸核糖:由乳清酸磷酸核糖转移酶催化乳清酸与PRPP反应,生成乳清酸核苷酸(orotidine-5′-monophosphate,OMP)。由PRPP水解供能。
(6)脱羧生成UMP:由OMP脱羧酶(omp decarboxylase)催化OMP脱羧生成UMP。
Jones等研究表明,在动物体内催化上述嘧啶合成的前三个酶,即CPS-Ⅱ,天冬氨酸氨基甲酰转移酶和二氢乳清酸酶,位于分子量约210kD的同一多肽链上,是一个多功能酶;因此更有利于以均匀的速度参与嘧啶核苷酸的合成。与此相类似,反应(5)和(6)的酶(乳清酸磷酸核糖转移酶和OMP脱羧酶)也位于同一条多肽链上。嘌呤核苷酸合成的反应(3)、(4)、(6),反应(7)和(8)及反应(10)和(11)也均为多功能酶。这些多功能酶的中间产物并不释放到介质中,而在连续的酶间移动,这种机制能加速多步反应的总速度,同时防止细胞中其它酶的破坏。
2.UTP和CTP的合成
三磷酸尿苷(UTP)的合成与三磷酸嘌呤核苷的合成相似。

三磷酸胞苷(CTP)由CTP合成酶(CTP synthetase)催化UTP加氨生成。(图8-9)动物体内,氨基由谷氨酰胺提供,在细菌则直接由NH3提供。此反应消耗1分子ATP。
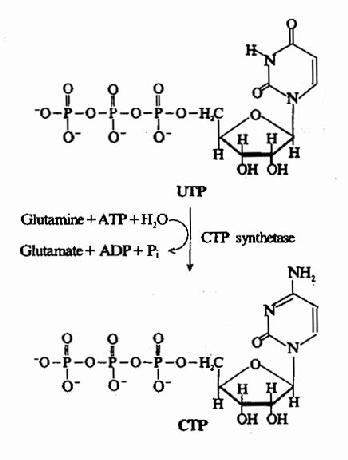
图8-9 由UTP合成CTP
3.嘧啶核苷酸从头合成的调节
在细菌中,天冬氨酸氨基甲酰转移酶(ATCase)是嘧啶核苷酸从头合成的主要调节酶。在大肠杆菌中,ATCase受ATP的变构激活,而CTP为其变构抑制剂。而在许多细菌中、UTP是ATCase的主要变构抑制剂。
在动物细胞中,ATCase不是调节酶。嘧啶核苷酸合成主要由CPS-Ⅱ调控。UDP和UTP抑制其活性,而ATP和PRPP为其激活剂。第二水平的调节是OMP脱羧酶,UMP和CMP为其竞争抑制剂。(图8-10)
此外,OMP的生成受PRPP的影响。
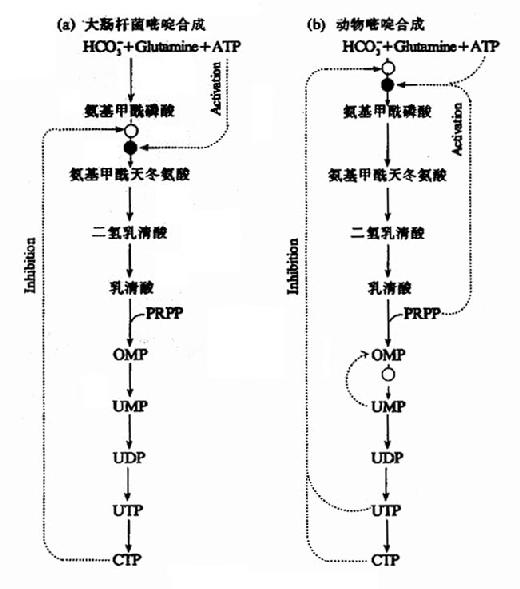
图8-10 嘧啶合成的调节网
4.乳清酸尿症(Orotic aciduria)
乳清酸尿症是一种遗传性疾病,主要表现为尿中排出大量乳清酸、生长迟缓和重度贫血。是由于催化嘧啶核苷酸从头合成反应(5)和(6)的双功能酶的缺陷所致。临床用尿嘧啶或胞嘧啶治疗。尿嘧啶经磷酸化可生成UMP,抑制CPSⅡ活性,从而抑制嘧啶核苷酸的从头合成。