�ڶ��ڡ��ļ���������������������
�ķ������Ҳ�ͣЪ�ؽ�����˳��ġ�Э�������������Ž���Ļ��������ʵ�ֱ�Ѫ���ܡ��ƶ�ѪҺѭ���ı�Ҫ��������ϸ��Ĥ���˷ܹ������Ǵ���������Ӧ��ʼ�����ء�������Ҫ�����������ǣ�����������������˷����Ժδ���Ϊʲô�����ĸ�ǻ���ܹ���Э�����������Ϊʲô����������ʼ�������������Ž����������ǿֱ������Ҫ�ش���Щ���⣬�����˽��ļ����������ԣ���Ҫ���ļ��˷ܺ��˷ܴ������������˷ܺʹ�������ϸ��Ĥ�������Ϊ�����ġ���ˣ����������ļ�ϸ�������������Ȼ���������������������ļ��˷ܺ��˷ܴ����Ĺ��ɺ��������塣
�ļ�ϸ�����������������ļ�ϸ��������ͬһ���͵ģ��������ǵ���֯ѧ�ص㡢�����������Լ������ϵ����𣬴��Եط�Ϊ�������ͣ������ļ�ϸ���ֱ�ʵ��һ����ְ�ܣ�������ϣ���������������һ������ͨ���ļ�ϸ���������ķ��������Ҽ������зḻ�ļ�ԭ��ά��ִ���������ܣ����ֳ�Ϊ����ϸ��������ϸ�������Զ��ز����������˷ܣ����������Զ������ԣ����������˷��ԣ������������̼������²����˷ܣ�Ҳ���д����˷ܵ����������ǣ�����Ӧ���������֯���Ƚϣ������Խϵ͡���һ����һЩ����ֻ��˵��ļ�ϸ�����������������ϵͳ��������Ҫ����Pϸ���Ͳ���Ұϸ�������dz��˾����˷��Ժʹ�����֮�⣬�������Զ������������˷ܵ��������ʳ�Ϊ����ϸ�������Ǻ���ԭ��ά��С����ȫȱ���������������ѻ���ɥʧ������һ��ϸ��λ�������ϵͳ�Ľ������Ȳ������������ܣ�Ҳû�������ԣ�ֻ�����˺ܵ͵Ĵ����ԣ��Ǵ���ϵͳ�еķ�����ϸ���������ϵͳ�������ڷ����˷ܺʹ����˷ܵ���֯�����ſ�����������Ի�����á�
���������ϵͳ����ɺͷֲ�����������ϵͳ�ɲ�ͬ���͵�����ֻ����ļ�ϸ������ɡ������ᡢ���ҽ��硢��������ĩ���ֿ�Ұ��ά����ͼ4-5����
�λ�����ķ�����ǻ�������Ӵ�����Ҫ����Pϸ������ϸ����Pϸ��������ϸ����λ�������IJ��֣�����ϸ��λ���ܱ߲��֣������������ԣ��������ǽ�Pϸ���Զ��������˷���������ķ�����
���ҽ��磺�ֳ�Ϊ���ҽ��������ķ�������֮����������֯�����ķ��˷ܴ������ҵ�ͨ�������ҽ�����Ҫ��������������������
��������λ���ķ��ͽ���֮�䣬���д����Ժ������ԡ�
�������൱�ڹ�ѧ���������ķ��ҽᣬ���д����ԣ��������ԡ�
��ϣ����λ�ڽ�����ϣ����֮�䣬���д����Ժ������ԡ�
���������ֳ�ϣ�����������֧���������������Ҽ���ڣ����Ҽ��Ĥ����ʼ��Ϊ������֧������֧��ϸ����;��֧�٣��ֲ��������ң�����֧�ʴ�״����֧�࣬�ֲ��������ң���������Ҫ���ֿ�Ұϸ����
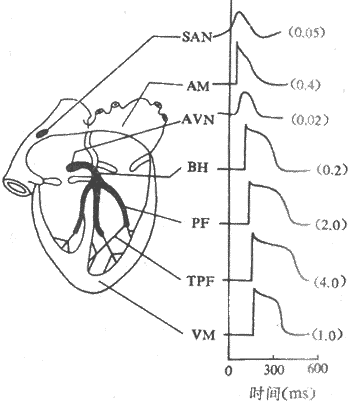
ͼ4-5 ����������ļ�ϸ���Ŀ�Ĥ��λ���˷ܴ����ٶ�
SAM���� AM���ķ��� AVN������ BH��ϣ���� PE������Ұ��ά
TPF��ĩ���ֿ�Ұ��ά VM�����Ҽ������ٶȵ�λm/s
�ֿ�Ұ��ά������������֧������֧�����ڷ�֧�ܶ࣬�γ���״���ܲ����������ҵ�����Ĥ�£�����ֱ������Ĥ�����ӣ�������ͨ���Ҽ�ϸ�������ӡ���������ĩ���ֿ�Ұ��ά�������ã��ǽ��ķ��������˷�Ѹ�ٴ������������ҡ�
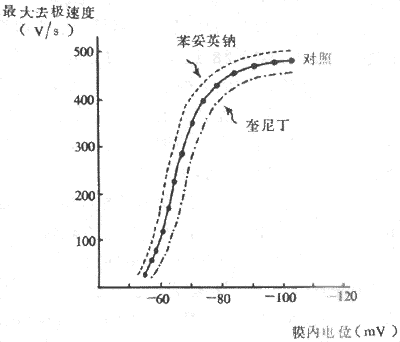
�����Ƿ�����ķ������������⣬���ۺܶࡣ60�����Janes�������ͷ��ҽ���֮�����������ֿ�Ұϸ�����ɵ��ķ����������ֱ��ǰ���С������������˷ܴ����ٶȱ�һ���ķ���Ϊ�졣���ǣ���20�������о�δ��֤ʵ�ķ�������̬�ṹ�ϲ�ͬ���ķ���������ϸ�����������֯��ɵ��ķ����������ڣ���һ���棬�о���������������ķ���ijЩ��λ������Բ��ǰ���ͽ��մ����ķ�����ά���з���һ�£��ṹ���룬����䴫���ٶȽ�������λ�ķ�������Щ�ķ��������ķ�����ǻ����������Բ�����γɵĿ�Ѩ���ָ�γɶ���״��Ϊ�죬�Ӷ��ڹ����Ϲ����˽����˷ܿ��ٴ��������ҽ��紦����ν���ƴ���ͨ·��preferential pathway�� ��
һ���ļ�ϸ�������������
���������ȣ��ļ�ϸ���Ŀ�Ĥ��λ�ڲ����Ϻ��γɻ�����Ҫ���ӵöࣻ������ˣ�������ͬ���͵��ļ�ϸ���Ŀ�Ĥ��λ��ͼ4-5�����������Ⱥͳ���ʱ�������ͬ�����Ҳ��κ��γɵ����ӻ���Ҳ��һ���IJ�𣻸����ļ�ϸ�����IJ�һ���ԣ��������˷ܵIJ����Լ��˷����������ഫ�������б��ֳ�������ɵ�ԭ��
��һ������ϸ���Ŀ�Ĥ��λ�����γɻ���
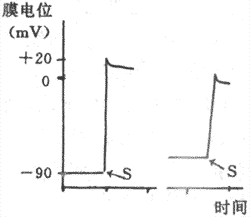
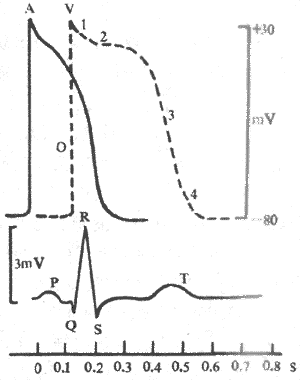
1����Ϣ��λ�Ͷ�����λ�˺Ͳ��鶯������Ҽ�ϸ��������ϸ��һ�����ھ�Ϣ״̬��Ĥ����ʼ���״̬��Ĥ�ڵ�λ��Ĥ���λԼ��90mV�������ߵĶ�����λ�����Բ�ͬ��������ϸ��������λ��ʱ�̺̣ܶ��������������룬�����ٶ���ȥ���ٶȼ�����ȣ���¼���߳���֧�ͽ�֧�����ԳƵļ��״�����Ҽ�ϸ��������λ����Ҫ�������ڸ������̱Ƚϸ��ӣ�����ʱ��ܳ���������λ��֧����֧�ܲ��Գơ�ͨ����0��1��2��3��4�����ֱַ�������Ҽ�ϸ��������λ�;�Ϣ��λ�ĸ���ʱ�ڡ�
��1��������ȥ�������̣����������ֳ�0�ڡ������˵������̼������£����Ҽ�ϸ�������˷ܣ�Ĥ�ڵ�λ�ɾ�Ϣ״̬�µ�-90mVѸ��������+30mV���ң�����Ĥ����ԭ�еļ���״̬���������ʼ�����ת�����ɶ�����λ����֧��������ܶ��ݣ���ռ1-2ms�����ҳ������Ⱥܴ�Ϊ120mV���ɼ������Ҽ�ϸ���ij����ٶȺܿ죬Ĥ��λ�����仯���ʿɴ�800-1000V/s��
��2���������̣�������ϸ�������ﵽ����֮��������ʼ�������������������̱Ƚϻ�����������λ�仯���ߵ���̬���γɻ��ƾ�����ͬ�������Σ�
1�ڸ������ڸ������ڣ������ֲ��ָ�����Ĥ�ڵ�λ��+30mVѸ���½���0mV���ң���1���ֳ�Ϊ���ٸ������ڣ�ռʱԼ10ms��0�ڳ�����1�ڸ���������ʱ�ڵ�Ĥ��λ�ı仯�ٶȶ��ܿ죬��¼ͼ���ϱ���Ϊ���״�������ļ�ϸ��ϰ���ϳ����������ֺϳ�Ϊ���λ��
2�ڸ�������1�ڸ���Ĥ�ڵ�λ�ﵽ0mV����֮�������̾ͱ�÷dz�������Ĥ�ڵ�λ������ͣ����0mV���ң�ϸ��Ĥ����ʵȵ�λ״̬����¼ͼ�αȽ�ƽ̹���ʸ���2���ֳ�Ϊƺ��ƽ̨�ڣ�����Լ100-150ms��������������λ����ʱ�䳤����Ҫԭ�������Ҽ�ϸ���Լ������ļ�ϸ���Ķ�����λ�����ڹ�����������ά����Ҫ������
3�ڸ�����2�ڸ��������У�����ʱ��Ľ�չ��Ĥ�ڵ�λ�Խ������ٶ���0mV���½�������Ϊ3�ڸ�����2�ں�3��֮��û�����ԵĽ��ޡ���3�ڣ�ϸ��Ĥ�����ٶȼӿ죬Ĥ�ڵ�λ��0mV���ҽϿ���½���-90mV����ɸ��������̣���3���ֳ�Ϊ���ٸ���ĩ�ڣ�ռʱԼ100-150ms ��
4�ڣ�4����Ĥ������ϡ�Ĥ��λ�ָ����ʱ�ڡ������Ҽ�ϸ��������������ϸ����4����Ĥ��λ�ȶ��ھ�Ϣ��λˮƽ����ˣ�4���ֿɳ�Ϊ��Ϣ�ڡ�
2���γɻ����������һ����������ϸ��Ĥ������ȷֲ����γɵ�Ũ���ݶȣ�Ũ�Ȳ����4-1����������Ӧ���Ӿ�����ʱ���ŵ�ϸ��Ĥ����������ͨ���Ŀ�Ĥ��ɢ�����ļ�ϸ����Ĥ��λ�γɵ���Ҫ������ֻ�������ļ�ϸ��Ĥ�Ͼ�����Ŀ�϶������ͨ������Ĥ��λ�γɻ������漰��������Զ�ȹ�����Ҫ���ӵöࡣ�ڵ�����ѧ�У������ķ�������������Ĥ����������������������������������������������������������������������������������������������Ĥ�ڵ�λ����ת������ʹĤ�����������������Ĥ�ڵ�λ��������ת������ʹĤ������
��4-1 �ļ�ϸ���и�����Ҫ���ӵ�Ũ�ȼ�ƽ���λֵ
| �� �� | Ũ�ȣ�mmo1/L�� ϸ����Һ ϸ����Һ | ��/���ֵ | ƽ���λ��mV�� (��Nernst��ʽ����) | |
| Na+ | 30 | 140 | 1:4,6 | +41 |
| K+ | 140 | 4 | 35:1 | -94 |
| Ca2+ | 10-4 | 2 | 1:20,000 | +132 |
| CI- | 30 | 104 | 1:3.5 | -33 |
�����ӿ�Ĥ��ɢ֮�⣬��ϸ�������ӱ���ʵ�ֵ���������ת�˺����ӽ��������ļ�ϸ������Ҳռ����Ҫ��λ��
���Ҽ�ϸ����Ϣ��λ���γɻ������������ͬ��Ҳ����˵�����ܼ�Ĥ���������������Ӷ�������Ũ���ݶȣ�����Ϣ״̬�¼�Ĥ��K+��ͨ�Խϸߣ������������ӵ�ͨ�Ժܵͣ���ˣ�K+˳��Ũ���ݶ���Ĥ����Ĥ����ɢ���ﵽ��ƽ���λ���Ǿ�Ϣ��λ����Ҫ��Դ��
��Ĥ��ͨ���Ĵ������ź�Ĥ����Ũ���ݶȼ���λ�ݶȵ������Ӷ�����Na+���������������Ҽ�ϸ��0��ȥ���γɵ�ԭ��һ��������ȥ�����̽��з����Ϳ��Կ������������һ�����������̼������£��������ֵ�ѹ�ſ�ʽNa+ͨ�����ź�����Na+��������ɼ�Ĥ����ȥ������Ĥ��λ����ֵ�½�������Ĥ��λ�ɾ�Ϣˮƽ��Ĥ��-90mV��ȥ�������е�λˮƽ��Ĥ��-70mV��ʱ��Ĥ��Na+ͨ�����Ÿ����������ӣ�����������Na+�������ο��ڶ��£�������Na+˳��Ũ���ݶȺ͵�λ�ݶ���Ĥ����ٽ���Ĥ�ڣ���һ��ʹĤȥ������Ĥ�ڵ�λ��������ת��������0��ȥ����Na+ͨ����һ�ֿ�ͨ����������������ŵ��ٶȺܿ죬���Ҽ����ܿ��ʧ���Ĥ������һ���̶ȣ�omV���ң�ʱ��Na+ͨ���Ϳ�ʼʧ����رգ������ֹNa+�ļ�����������Na+ͨ���ɱ������TTX������ϡ�����Na+ͨ�������ٶȷdz�֮�죬����������ѭ�����֣���������Ҽ�ϸ��0��ȥ���ٶȺܿ죬������λ��֧�dz����͵�ԭ������Ϊ��ˣ��ӵ����������ϣ������Ǹ���0�ڳ��������ʣ������Ҽ�ϸ�����Լ�����ͬ���������ļ�ϸ������Ϊ�췴Ӧϸ�����䶯����λ��Ϊ�췴Ӧ��λ�����������Ժ�Ҫ���ܵ�����Ӧϸ��������Ӧ��λ��
����1������0�ڳ���֮����ֵĿ��ٶ����ݵĸ����ڣ���ʱ����ͨ���Ѿ�ʧ�ͬʱ����һ��һ�������������Ito�����Ӷ�ʹĤѸ�ٸ�����ƽ̨�ڵ�λˮƽ��0��-20mV��������Ito�����ӳɷ֣�70�������Ϊ��Cl-����Cl-��������������������Ito�ɱ����һ�狀�4-������व�K+ͨ�����ͼ�����ϵ��о����ϣ���ΪK+����Ito����Ҫ���ӳɷ֡�Ҳ����˵����K+���ص�һ������������Ƕ�����λ���ڿ��ٸ�������Ҫԭ��Ŀǰ��Ito��ͨ�������в�ʮ�����������������ʾ��Ĥ������ϸ����Ca2+������ʹIto��ͨ�����
ƽ̨�ڳ��ڣ�Ĥ��λ�ȶ���0mV���ң����ŷdz������ظ�����Ĥ��λ����������������ƽ̨��ͬʱ���������������������ڣ����ڣ����ֵ����������ƽ��״̬�����������������������������ǿ���ܺ͵Ľ���dz���һ����ʱ�����ƶ�����ǿ�ġ������������������Ĥ��λ��������Ĥ�ڸ�����ת������ѹǯ�о���������������Ҽ��ȿ췴Ӧϸ����ƽ̨����������������K+Я���ģ���Ik1������Ϣ״̬�£�K+ͨ����ͨ�Ժܸߣ���0�ڳ��������У�K+��ͨ�������½���K+���������٣����������ʱ��K+��ͨ�Բ����������ָ�����Ϣ״̬�µ����ָ�ˮƽ�����Ǽ��仺���ء����ֵػָ���K+����Ҳ���ɳ��ڵĵ�ˮƽ���������ӣ�ͼ4-6����ƽ̨��������������Ҫ����Ca2+(�Լ�Na+)���صġ��Ѿ�֤�����ļ�ϸ��Ĥ����һ�ֵ�ѹ�ſ�ʽ����Ca2+ͨ������Ĥ������-40mVʱ�����Ca2+˳��Ũ���ݶ���Ĥ�ڻ�����ɢ�Ӷ�������ʹĤ���������ͬʱ����������K+����������ʹĤ����������ƽ̨�����ڣ�Ca2+��������K+�����������صĿ�Ĥ�����ʱ��ȣ�Ĥ��λ�ȶ���1�ڸ������ﵽ�ĵ�λˮƽ������ʱ�����ƣ�Ca2+ͨ����ʧ�K+���������ӣ���������Ĥ�ľ�������������ӣ�Ĥ�ڵ�λ�������½����γ�ƽ̨�����ڡ��˺�Ca2+ͨ����ȫʧ�������������ֹ������K+����һ����ǿ��ƽ̨������Ϊ����3�ڣ�Ĥ��λ�Ͽ�ػص���Ϣˮƽ����ɸ��������̡�
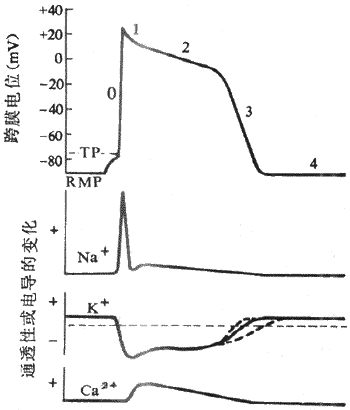
ͼ4-6 ���Ҽ�ϸ����Ĥ��λ�����γɵ����ӻ���
RMP����ϢĤ��λ TP ���е�λ
��Ĥ����Ca2+ͨ���������Ҽ�ϸ���������ļ�ϸ������Ҫ�����������о��������ٴ�һ���ļ�ϸ����������ԣ����Ǵӵ���ͨ�����ԣ���Ca2+ͨ���ļ��ʧ��Լ��ٸ�������ʱ�����Naͨ��Ҫ������ Ca2+ͨ����Ĥ��Ca2+��������ʼ����ƽ������ʱ��Ҳ�ϳ��������Ӧ��Ϊ��ͨ����������������������ͨ��Ҳ�ǵ�ѹ�ſ�ʽ�ģ�������ͨ�����е�λˮƽ��-50��-35mV�����ڿ�Naͨ����-70��-55mV����������ijЩ�������ص������Ժͷ�Ӧ�Բ�ͬ�ڿ�ͨ�����ɱ�Mn2+�Ͷ���Ca2+��ϼ������첩����D-600�ȣ�����ϣ������ڿ�����Ͽ�ͨ���ĺ����ϸ��Ĥ�ij����ͼ���״̬��Ĥ�ڵ�λ-50Mv���ң�ȴ�������С������ļ�ϸ���ļ�Ĥ�϶�����������ͨ�����ɴ��γɵĿ�Ĥ���������Ǿ����ļ�ϸ�����Լ����Ҽ��ȿ췴Ӧϸ��������λƽ̨�ڵ�����Ҫ������������֮һ��
ƽ̨��֮��Ĥ�ĸ������٣����ʱCa2+ͨ���Ѿ�ʧ���ƽ̨���Ѿ����������K+��������ʱ������������ơ���ԭ���ǣ�3�ڵĸ���K+���������Եģ�K+��������ʹĤ�ڵ�λ����ת������Ĥ�ڵ�λԽ����K+������Խ���ߡ��������������̣�����Ĥ�ĸ���Խ��Խ�죬ֱ����������ɡ�
��4���ڣ����Ҽ�ϸ��Ĥ��λ�������ȶ��ھ�Ϣ��λˮƽ�����ǣ����ӵĿ�Ĥת����Ȼ�ڻ�Ծ���С���Ϊ��������λ�ڼ���Na+��Ca2+����ϸ���ڣ���K+������ϸ������ˣ�ֻ�д�ϸ�����ų������Na+��Ca2+��������K+���ָܻ�ϸ���������ӵ�����Ũ���ݶȣ������ļ�ϸ���������˷��ԡ���������ת��������Ũ���ݶȽ��е�����ת�˹��̡��������һ����ͨ����Ĥ��Na+-K+�õ����ã���Na+�����˺�K+�����˻��������γ�Na+-K+ת�ˣ�ͬʱʵ��Na+��K+������ת�ˡ���������ת��Ca2+��ת�˻��ƣ���û����ȫŪ�����Ŀǰ�����������Ϊ��Ca2+����Ũ���ݶȵ���������Na+��˳Ũ�ȵ���������Ͻ��еġ��γ�Na+-Ca2+������Ca2+����������ת������Na+ ��������Ũ���ݶ��ṩ�����ģ�����Na+������Ũ���ݶȵ�ά��������Na+-K+�ö�ʵ�ֵģ���ˣ�Ca2+����ת��Ҳ����Na+-K+���ṩ�����ġ���4�ڿ�ʼ��Ĥ����������ת�˹��ܼ�ǿ��ϸ����������Ũ���ݶȵ��Իָ����ܵ���������ʱת�˹�������Ŀ�Ĥ�����ĵ����������ȣ���ˣ�Ĥ��λ����Ӱ�����ά���ȶ���
����������ϸ���Ŀ�Ĥ��λ�����γɻ���
��û�������̼�ʱ������ϸ�����ܲ���������λ���������̼������£�����һ�ζ�����λ�������ζ�����λ֮��Ĥ��λ���ȶ�����ġ���������ϸ������������λ3�ڸ���δ�ڴﵽ���ֵ���������λ��֮��4�ڵ�Ĥ��λ�����ȶ�����һˮƽ������������ʼ�Զ��������������е�λ�������˷ܣ�������һ��������λ�����������ܶ���ʼ��������λ�Ͳ��ϵز�����������4�ڵ������Զ��������̣�������ʱ����������ص㣬������ٶ�Զ��0�ڳ�����������ͬ���͵�����ϸ��4�ڳ����ٶȲβһ����ͬ������ϸ��4�ڳ����ٶȱȽϺ㶨������4���Զ����������4�ڻ����������������ڳ�������������ϸ�������Զ��������˷ܵĻ�����
����ϸ��Ĥ�����Ŀ�Ĥ�����Ļ������ɿɷ�������ϸ��4���Զ������γɵĻ��ơ������Ʋ⣬����ϸ�����ھ��������ʹĤ������3�ڣ��������λ����4�����ֳ���һ������ǿ�ľ�����������Ӷ�ʹĤ������λ�����ӣ�Ĥ�����������ֽ����Ծ���������IJ��������������ֿ��ܵ�ԭ���������������ǿ���������������˥�ˣ��������С���ͬ���͵�����ϸ����4���Զ��������������ֽ����Ծ�������������𣬵����ɾ�����������������ķ�������ӱ��ʲ�����ȫ��ͬ��
1���ֿ�Ұϸ���ֿ�Ұϸ����һ�ֿ췴Ӧ����ϸ������Ϊһ�ֿ췴Ӧ��ϸ�������Ķ�����λ����̬�����Ҽ�ϸ�����ƣ����������ӻ���Ҳ������ͬ��
�����ֿ�Ұϸ��4���Զ������γɵĻ��ƣ�80����о����ϱ��������ֿ�Ұϸ�������Ÿ����Ľ��У�����Ĥ����������K+������˥������ͬʱ��Ĥ��λ4�ڿɼ�¼��һ����ʱ�����ƶ�����ǿ�����������If����ͼ4-7����Ifͨ���ڶ�����λ3�ڸ�����λ�60mV���ҿ�ʼ������ţ��伤��̶����Ÿ����Ľ��С�Ĥ�ڸ����Ե����Ӷ����ӣ�����100mV���Ҿͳ�ּ����ˣ�����������ֳ�ʱ����������ǿ��Ĥ�ij����̶����Ҳ��ʱ������ӣ�һ���ﵽ�е�λˮƽ�����ֲ�����һ�ζ�����λ�����ͬʱ���������������Ĥ������-50mV������ͨ��ʧ�����ֹ���ɼ���������λ�ĸ�����Ĥ��λ��������������������������ͷ�չ�����أ���������IJ�������ǿ����Ĥ�Ľ����Գ�������Ĥ�ij���һ����������һ�ζ�����λ��һ�����ַ�������ֹ���������������һ�����Ĺ���������ϸ�������ҡ������������ҡ���չ���֡����ҡ����Ƶģ��ɴ˿�������Ϊʲô����ϸ���ܹ��Զ��ء����ϵز����������˷ܡ�
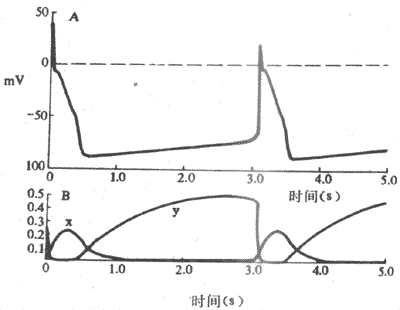
ͼ4-7 �ֿ�Ұϸ������
A����Ĥ��λ B���ɧ�բ�ſ��Ƶ�Ik˥���Լ��ɧ�բ�ſ��Ƶ�If���������γ���λ�е���Թ�ϵ
����4�����������ͨ����Ϊ����������Ҫ���ӳɷ�ΪNa+ ����Ҳ��K+���롣����ʹ����ּ����Ĥ��λΪ-100mV�������Ϊ�������������������һ�ֱ�Ĥ�ij���������ķ�������������Ҫ����Na+������������־����ΪIf��If��ͨ������Na+ͨ��������ͬ�ڿ�Na+ͨ����������ĵ�ѹˮƽ��ͬ��If�ɱ�藍�Cs������ϣ������ȴ�����������Ŀǰ������If����ͨ�����о������������ɲ��ܳ�����Խ��͵��ɵ㣬��If�Ľ�һ���о����ܵ��ļ�������ѧ���ǵĸ߶ȹ�ע��
2����ϸ���Ŀ�Ĥ��λ�����γɻ��� �Ậ�зḻ������ϸ����������λ������������Ե�4���Զ�������������һ������Ӧ����ϸ�������Ĥ��λ�������ͬ�����Ҽ��췴Ӧϸ�����ֿ�Ұ�췴Ӧ����ϸ��������������ϸ���������λ��-70mV�����е�λ��-40mV�������ڣ���λ�������ֿ�Ұϸ������0�ڳ�������ʱ��Ĥ�ڵ�λΪ0mV���ң����������Եļ�����ת������������ȣ�70mV��С���ֿ�Ұϸ����Ϊ120mV��,��0�ڳ���ʱ�̣�7ms���ң�ȴ�ֱȺ��ߣ�1-2ms�����öࡣԭ������ϸ��0�ڳ����ٶȣ�Լ10V/s�����������ֿ�Ұϸ����200-1000V/s������ˣ�������λ��֧Զ���������ô���ͣ���û�����Եĸ���1�ں�ƽ̨�ڣ���4���Զ������ٶȣ�Լ0.1V/s��ȴ���ֿ�Ұϸ����Լ0.02V/s��Ҫ��,��¼��������ϸ��4��Ĥ��λ�仯��б�ʴ����ֿ�Ұϸ��.ͼ4-8��ʾ���Ҽ��췴Ӧϸ������ϸ����Ĥ��λ�仯�IJ��
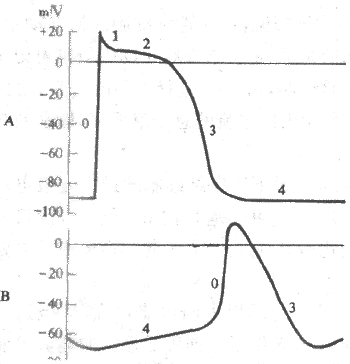
ͼ4-8 ���Ҽ���A�����ᣨB��ϸ����Ĥ��λ�ıȽ�
��ϸ����ֱ����С�����е������о���һ�����ѡ�ֱ��70������ڣ��ſ�ʼ����С�걾�ϲ��õ�ѹǯ���������Ĥ�����������˶����о�����Ŀǰ��δ�ܳ�ֲ������Ŀ�Ĥ��λ��������4�����������ӻ�����ѧ���ǹ۲쵽����ϸ��0�ڳ�������ϸ����Na+Ũ�ȵ�Ӱ�죬�Ժ���ܲ����У��෴������ϸ����Ca2+Ũ�ȵ�����Ӱ�죬���ɱ����Ƹ�ͨ����ҩ������ӣ����첫����D-600��Mn2+�ȣ�����ϡ��ݴ˿�����Ϊ��������ϸ��������λ0�ڳ����������������Ca2+���صġ����������������Ϊ�ڶ����������������췴Ӧϸ�������Ҽ����ķ������ֿ�Ұϸ����0�ڳ����Ŀ�Na+������Ϊ��һ����������������е��о����ϣ��ɽ���ϸ��������λ���γɹ����������£���Ĥ��λ�������λ�Զ��������е�λˮƽʱ������Ĥ�ϸ�ͨ��������Ca2+��������Ica��������0�ڳ��������ͨ����ʧ�Ca2+������Ӧ���٣���һ���棬�ڸ������ڣ���һ��K+ͨ�����������K+��������Ik����Ca2+���������ٺ�K+�����������ӣ�Ĥ���������ɡ�����ͨ�������ơ���Ca2+����������Ļ���0�ڳ���������ϸ��������λ����Ҫ��������ˣ���Ӧ��Ϊ����Ӧϸ��������Ӧ��λ����������ǰ�����Ҽ��ȿ췴Ӧϸ���Ϳ췴Ӧ��λ��
��ϸ����4���Զ�����Ҳ����ʱ��������ľ�������������𣬵��乹�ɳɷֱȽϸ��ӣ��Ǽ��ֿ�Ĥ�������Ļ�ϡ�Ŀǰ��֪������ϸ��4�ڿ��Լ�¼������Ĥ����������һ����������������������������������ϸ������������õĴ�С�Լ������õ�ʱ��������ͬ��
��1��Ikͨ���ļ��������ǿ����ɵ�K+���������ǵ�����ϸ��������ԭ��Ikͨ����Ĥ������-40mVʱ�㿪ʼ��ʧ�K+������˽������٣�����Ĥ������������Ӷ��γ�4�ڳ�����Ŀǰ��Ϊ������Ikͨ����ʱ����������ʧ������ɵ�K+����������˥��������ϸ��4���Զ�����������Ҫ�����ӻ�����ͼ4-9����
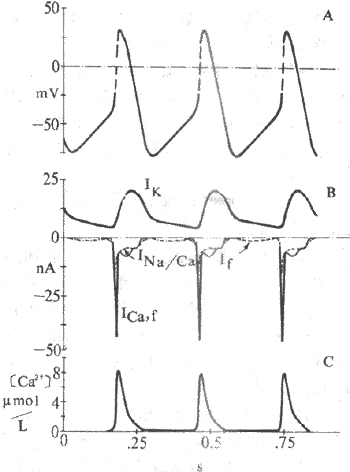
ͼ4-9 �ᶯ����λ����λ�����ӻ���
A����Ĥ��λ B.ԽĤ��λC. ����Ca2+Ũ�ȱ�ʾ������λ��֧��Ica,
f���ɣ���λ ��Ik��If��Ina/Ca����
(2)If��If��һ�ֽ�������ǿ���������ӣ���ҪΪNa+���������ֿ�Ұϸ����У�If���ż���Ҫ�����ã���Ik˥�������ú�С�����ǡ�෴����ϸ��4����Ҳ�ɼ�¼��If �����������������ò���Ik˥����ʵ��֤������Cs2+ѡ�������If�����Է���Ƶ�ʽ���ȼ��٣��Լ�����ϸ��4�ھ�����������ܷ�ֵ���ԣ�Ik˥����If�����������õı���Ϊ6��1��
Ifͨ��������λΪ-100mV���ң��������������ϸ���������λΪ-70mV�������ֵ�λˮƽ�£�Ifͨ���ļ���ʮ�ֻ������������If����4�ڳ����������������ò����ԭ������ϸ������������ʱ��If����ܳ�Ϊ�����е���Ҫ�ɷ֡�
��3�����⣬��ϸ��4���л�����һ�ַ������ԵĻ��������������Ĥ������-60mVʱ������ɼ��������Զ��������̵ĺ�1/3�ڼ�������á����ֻ����������������������Na+-Ca2+�����Ľ����Na+-Ca2+����ʱ���ļ�ϸ���ų�һ��Ca2+������3��Na+����/��ϸ�������֮��Ϊ2��3���γ������������
�ļ�ϸ�������� ���˰��չ��ܺ͵��������Խ��ļ�ϸ����Ϊ����ϸ��������ϸ��֮�⣬�����Ը������������������ر��Ƕ�����λ0�ڳ������ٶȣ����ļ�ϸ����Ϊ�췴Ӧϸ��������Ӧϸ�����䶯����λ��Ӧ��Ϊ�췴Ӧ��λ������Ӧ��λ��Ȼ���ٽ���������ԣ��ɽ��ļ�ϸ����Ϊ�����������ͣ�
�췴Ӧ������ϸ���������ķ���ϸ�������Ҽ�ϸ����
�췴Ӧ����ϸ�����ֿ�Ұ����ϸ����
����Ӧ����ϸ����������ϸ�����Լ��������ͽ�ϣ��������ϸ����
����Ӧ������ϸ��������ϸ����
�ļ�ϸ���Ŀ�Ĥ������ �ɶ�������ͨ��ϸ��Ĥ�ϵ������Ի������������ͨ���Լ������ӽ���ת���γɵĿ�Ĥ�����˶������ļ�ϸ����Ĥ��λ�γɵĻ������ļ�ϸ����Ĥ�������������ڶ࣬���ʺܸ��ӣ�������ȡ�������չ��������ʵ���о��ж���Щ����ͨ������������ʶ�Ƚϻ��ң�����ǰ��ì�ܣ��ʽ�Ŀǰ�ȽϿ϶�����Ҫ������������ͨ�������ơ����ڵIJ�λ�����ù��ɽ������£�
1������������
��1��INa����Ϊ����Na+���������ڿ췴Ӧϸ����������췴Ӧϸ��0�ڳ���������������Ƶ�һ��������������ӻ�����
��2��Isi����Ϊ�������������Ҳ�Ƶڶ������������ȥ��Ϊ����һ������ͨ�����ƵĻ���Ca2+�����ʱ�־ΪICa���书���ǹ��ɿ췴Ӧϸ����ƽ̨�ں�����Ӧϸ���ij����ڡ��½����о�����ںܴ�̶��������˶�Ica����ʶ��Ŀǰ��Ϊ���ڶ�������������ǵ�һ��Ca2+�������������Ը����������������ɣ���һ��ֳ�Ica.f����������ֱ��Is1.2��Ica.s�����Т�Ica.f��һ�ֿ���Ca2+������ͨ�������ʧ����ٶ�Զ����������Ϊ��Ҫ��öࡣ���ں��ڿ췴Ӧϸ��Na+��������֣���ͬ�γɶ�����λ��֧���϶Σ�����ƽ̨�ڵ����ú�С����������Ӧϸ�����������ӻ������Թ���ϸ�����ԣ�������Ca2+�������ͷ�˧ϸ����Ca2+������������ģ���ȻIca.f�ļ���ʮ��Ѹ�٣���ôCa2+�ͷŵĴ����м��˷ܣ����������������ٶȶ���������Ϊ��Ҫ��öࣻ�ڵ������Ica.s����һ�ֽ�Ica.f����������Ca2+������Ҫ������ά�ֿ췴Ӧϸ��ƽ̨�ڣ���Is1.2�Ƶڶ���������ڶ���֣������ӱ��ʲ��������������Na+-Ca2+�����������������Ҳ��ΪINa.Ca����ƽ̨�������ã�����Ӧ����ϸ�������Զ���������Ҳ���������á�
(3)If������������ķ���������������������Ҫ��Na+Я��������������ϸ��4�ڡ�If�Ǿ����ֿ�Ұ�췴Ӧ���������Ҫ��������������������Ӧ����ϸ����У������ò���Ik˥����
2������������
��1��Ikl�������ڿ췴Ӧϸ�����Ǿ����췴Ӧ����ϸ����Ϣ��λ�������������ڸ���2�ں�3�������ã�
��2��Ik���������������Ҫ��K+Я������Ҳ��Na2+�μӣ����ǵ�����K+�������ֳ�Ix���ڿ췴Ӧϸ������3������Ҫ���ã����ֳ�Ϊ�������������Ҳ�������ֿ�Ұϸ���������λ��IxҲ����������Ӧ����ϸ������ʹĤ������4���ڳʽ�����˥�������γ�4�ڳ�������Ҫ���ӻ�����
��3��Ito���ǿ췴Ӧϸ��1�ڸ����������������ӳɷ���ҪΪK+��Ҳ��Na+���롣
3����ѧ�ſ�����ͨ�� �������֤ʵ�����ļ�ϸ��Ĥ�ϣ����˵�ѹ�ſ�ͨ���⣬�����ڻ�ѧ�ſ�����ͨ�������ǵ����ò�������ά�ֺͲ���������Ĥ��λ����Ϣ��λ�Ͷ�����λ�����������ļ�ϸ����ĵ����Լ��쳣���ĵ����������ر���Ҫ�����塣��Ҫ�Ļ�ѧ�ſ�����ͨ�����������֣�
��1������������Ƶ�K+ͨ����Ik-ACh������������ΪACh�������Ikl�����귢����ͨ��G�����������һ��������������ѧ���Ժ�����ѧ�����Ͼ�����ͬ��K+ͨ����
��2��ATP������K+ͨ����Ik-ATP����ATP�����ò����Ƿֽ�ܼ����ͨ��������ά�ִ�ͨ������������´��ڹر�״̬�����ļ�ϸ����ATP�����ٽ�ˮƽ����ʱ�����ļ�ȱѪʱ�������������K+ͨ�����š�����K+��©����ȱѪ�ļ�ϸ���ֲ��ض�����������շ�����ʧ����������ҩ�����ϴ�ͨ����
��3��INa�Ck��Ca2+ͨ������ϸ����Ca2+����ʱ�����һ�ַ�������������ͨ��������������Na+��K+���γ�һ����������������Iti����ʵ���������ijЩ����£��ֿ�Ұϸ���ڶ�����λ������ɲ���һ�ֳ�����λ���ӳٺ������λ�����������е�λʱ�Ϳ����շ���һ���µĶ�����λ���γ���λ������Iti�����ӳٺ������λ�����ӻ��������ػ��ж���ϸ�����K+���Na+���Լ������������Ӱ��ȿ�����ϸ����Ca2+�����ɵ����أ������շ����ǿIti���ӳٺ������λ��
�����ļ��ĵ���������
�ļ���֯�����˷��ԡ������ԡ������Ժ������������������ԡ��ļ�����������ָ�ļ��ܹ��ڼ�Ĥ������λ�Ĵ����²���������Ӧ�����ԣ�����������������֮������ﻯѧ������������ӦΪ�����ģ����ļ���һ�ֻ�е���ԡ��˷��ԡ������Ժʹ����ԣ������Լ�Ĥ�������Ϊ�����ģ����ֳ�Ϊ���������ԡ��ļ���֯����Щ�������Թ�ͬ����������Ļ��
��һ���ļ����˷���
�����ļ�ϸ���������˷��ԣ����������ܵ��̼�ʱ�����˷ܵ������������ļ����˷��ԣ�ͬ�����Բ��ô̼�����ֵ��ָ�꣬��ֵ���ʾ�˷��Եͣ���ֵС��ʾ�˷��Ըߡ�
1��������Ӱ���˷��Ե����� �ӹ����˷ܲ������̵������п�֪���˷ܵIJ���������Ϣ��λȥ�������е�λˮƽ�Լ�Na+ͨ�����Կ췴Ӧ��ϸ��Ϊ�����ļ��������������ڣ���������������ط����仯ʱ���˷��Խ���֮�����ı䡣
��1����Ϣ��λˮƽ����Ϣ��λ��������ϸ������Ϊ�����λ������ֵ����ʱ�������е�λ�IJ��ͼӴ������˷�����Ĵ̼���ֵ������Ϊ�˷��Խ��͡���֮����Ϣ��λ����ֵ����ʱ�����е�λ�IJ����С������Ĵ̼���ֵ���٣��˷������ߡ�
��2���е�λˮƽ���е�λˮƽ���ƣ���;�Ϣ��λ֮��IJ�����������˷�����Ĵ̼���ֵ�����˷��Խ��͡���֮��Ȼ��
��Ϣ��λˮƽ�ͣ����е�λˮƽ�ĸı䣬���ܹ�Ӱ���˷��ԣ��������࣬�Ծ�Ϣ��λˮƽ�ı�Ϊ�����ԭ��
��3��Na+ͨ������״�������˷ܵIJ���ʱ��������Na+ͨ���ܹ���������Ϊǰ�ᡣ��ʵ�ϣ�Na+ͨ��������ʼ�մ������ֿɱ������״̬�����ɱ���Ϊ���ʧ��ͱ������ֹ���״̬����Na+ͨ������������һ��״̬����ȡ���ڵ�ʱ��Ĥ��λ�Լ��йص�ʱ����̡������˵��Na+ͨ���Ļ�ǵ�ѹ�����Ժ�ʱ�������Եġ���Ĥ��λ����������Ϣ��λˮƽ-90mVʱ��Na+ͨ�����ڱ���״̬������״̬�£�Na+ͨ������˫�����ԣ�һ���棬Na+ͨ���ǹرյģ���һ���棬��Ĥ��λ�ɾ�Ϣˮƽȥ�������е�λˮƽ��Ĥ��-70mV��ʱ���Ϳ��Ա����Na+ͨ��Ѹ�ٿ��ţ�Na+������Կ��ٿ�Ĥ������Na+ͨ������������Ѹ��ʧ���ʱͨ���رգ�Na+����Ѹ����ֹ��Na+ͨ���ļ����ʧ����DZȽϿ��ٵĹ��̣�ǰ����1ms �ڣ�����Լ�ڼ����뵽10ms�ڼ�����ɡ�����ʧ��״̬��Na+ͨ������������Na+�Ŀ�Ĥ��ɢ�����Ҳ��ܱ��ٴμ��ֻ����Ĥ��λ�ָ�����Ϣ��λˮƽʱ��Na+ͨ�������»ָ�������״̬�����ָ����˷ܵ�������������̳�Ϊ������Ͽɼ���Na+ͨ���Ƿ���״̬���Ǹ��ļ�ϸ����ʱ�Ƿ�����˷��Ե�ǰ���������ϢĤ��λˮƽ���Ǿ���Na+ͨ���ܷ��ڻ��ܷ����״̬�Ĺؼ���Na+ͨ��������������״�����Խ����й��ļ�ϸ���˷��Ե�һЩ�������磬��Ĥ��λ��������Ϣˮƽ��-90mV��ȥ�������е�λˮƽ��-70mV��ʱ��Na+ͨ����������ֶ�����λ���������Ϣ״����Ĥ��λΪ-50mV���ң�����Ĥ���ڳ����ͼ���״̬ʱ���Ͳ�������Na+ͨ���������Ϊ�˷��Ե�ɥʧ������Na+ͨ����������״̬��ʵ���Լ�Ĥ��λ�����Ӱ��Na+ͨ����״�����⣬Ŀǰ��δ���ײ�����
2��һ���˷ܹ������˷��Ե������Ա仯�ļ�ϸ��ÿ����һ���˷ܣ���Ĥ��λ������һϵ���й��ɵı仯��Ĥͨ���ɱ���״̬�������ʧ�����ȹ��̣��˷���Ҳ��֮������Ӧ�������Ըı䡣�˷��Ե����������Ա仯��Ӱ�����ļ�ϸ�����ظ��̼��ķ�Ӧ���������ļ���������Ӧ���˷ܵIJ������������̾�����Ҫ���á����Ҽ�ϸ��һ���˷ܹ����У����˷��Եı仯�ɷ����¼���ʱ�ڣ�ͼ4-10����
��1����Ч��Ӧ�ڣ��ļ�ϸ������һ���˷ܺ��ɶ�����λ��ȥ���ʼ������3��Ĥ�ڵ�λ�ﵽԼ-55mV��һ��ʱ���ڣ�������ܵ��ڶ����̼������۴̼��ж�ǿ����Ĥ�������һ�������κγ̶ȵ�ȥ������Ĥ�ڵ�λ��-55mV�����ָ���Լ-60mV��һ��ʱ���ڣ��������Ĵ̼����㹻��ǿ�ȣ���Ĥ�ɷ����ֲ��IJ���ȥ�������������������������˷ܣ�������λ�����ļ�ϸ���˷ܺ��������ٲ����ڶ����˷ܵ����ԣ���Ϊ��Ӧ�ԣ���Ӧ�Ա���Ϊ����ġ����ݵ��˷���ȱʧ���½����ļ�ϸ��һ���˷ܹ����У���0�ڿ�ʼ��3��Ĥ�ڵ�λ�ָ���-60mV��һ�β����ٲ���������λ��ʱ�ڣ���Ϊ��Ч��Ӧ�ڡ���ԭ�������ʱ����Ĥ��λ����ֵ̫�ͣ�Na+ͨ����ȫʧ�ǰһ�Σ�����ոտ�ʼ�����һ�Σ�������ԶԶû�лָ������Ա�����ı���״̬��Ե�ʡ�
��2����Բ�Ӧ�ڣ�����Ч��Ӧ����ϣ�Ĥ�ڵ�λԼ-60mV������������������ɣ�Լ-80mV��������ڼ䣬Ϊ��Բ�Ӧ�ڡ���һʱ���ڣ�ʩ�Ӹ��ļ�ϸ���Ը���������ֵ��ǿ�̼������������������˷ܡ�������Բ�Ӧ�ڵ�ԭ���ǣ�����Ĥ��λ����ֵ������Ч��Ӧ��ĩʱ��Ĥ��λ�����Ե��ھ�Ϣ��λ����ʱNa+ͨ����������俪��������δ�ָ����������ļ�ϸ�����˷��������Ч��Ӧ��ʱ�����ָ�������Ȼ���������������˷�����Ĵ̼���ֵ�������������������Ķ�����λ������ǰ�˷ܣ�0�ڵķ��Ⱥ��ٶȶ�������ΪС���˷ܵĴ���Ҳ�Ƚ��������⣬���ڴ���ǰһ��������λ��3�ڣ�����K+Ѹ�����������ƣ���ˣ��ڴ������²����Ķ�����λ����ʱ�̽϶̣�K+������ʹƽ̨�����̣�����Ӧ��Ҳ�϶̡�
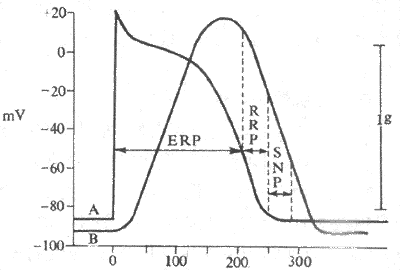
ͼ4-10 ���Ҽ�������λ�ڼ��˷��Եı仯�������е�����Ĺ�ϵ
A��������λ B����е����ERP����Ч��Ӧ��
RRP����Բ�Ӧ�� SNP��������
��3�������ڣ��ļ�ϸ������������Ĥ�ڵ�λ��-80mV�ָ���-90mV��һ��ʱ���ڣ�����Ĥ��λ�Ѿ������ָ����������ֵ�е��ھ�Ϣ��λ�����е�λˮƽ�IJ���С�����������ϸ�������˷�����Ĵ̼���ֵ������Ҫ�ͣ������˷��Ը����������ʳ�Ϊ�����ڡ���һ���棬��ʱNa+ͨ�������ϻָ����ɱ��������������״̬��������������Ȼû�лָ������������Ķ�����λ��0��ȥ���ķ��Ⱥ��ٶȣ��˷ܴ������ٶȶ���Ȼ����������
�������ϣ�Ĥ��λ�ָ�������Ϣˮƽ���˷���Ҳ�ָ�������
3���˷ܹ����У��˷��������Ա仯��������Ĺ�ϵϸ���ڷ���һ���˷ܹ����У��˷��Է��������Ա仯���������ͼ���֯��ͬ�����ԣ����ļ�ϸ������Ч��Ӧ���ر�һֱ��������е��Ӧ�������ڿ�ʼ֮����ˣ�ֻ�е���������֮���˷��Ա仯������Բ�Ӧ�ڣ����п������ܵ�ǿ�̼�����ʱ�����˷ܺ���������������ʼ����������֮�䣬�ļ�ϸ����������ڶ����˷ܺ�����������ص�ʹ���ļ����������������������ȫǿֱ������ʼ���������������ཻ��Ļ���Ӷ�ʹ������ѪҺ���ij�ӯ��ʱ�ڣ������ſ���ʵ�����Ѫ���ܡ�
����ʵ�����˵���ļ���֯����һ�ص㡣��������£��������ÿһ���˷ܴ������ķ��������Ҽ���ʱ�䣬����������ǰһ���˷ܵIJ�Ӧ���ս�֮����ˣ����������ܹ�������Ľ��ɶ��˷ܡ�����ijЩ����£������������Ч��Ӧ��֮���ܵ��˹��Ļ���֮��IJ������쳣�쳣�̼�����ɲ���һ����ǰ�˷ܣ�������ǰ�����������������ǰ�˷�Ҳ�����Լ�����Ч��Ӧ�ڣ�����������������ǰ�˷�֮���һ�����˷ܴ������Ҽ�ʱ����������������ǰ�˷ܵ���Ч��Ӧ���ڣ�����������������˷ܺ��������γ�һ�Ρ���ʧ��������ȵ�����һ������˷ܴ�������ʱ��������������������������һ����ǰ����֮����������һ�νϳ������������ڣ���Ϊ�����Լ�Ъ��ͼ4-11������֮���Żָ���Խ��ɡ�
�������ļ����Զ�������
��֯��ϸ���ܹ���û�������̼��������£��Զ��ط����������˷ܵ����ԣ���Ϊ�Զ������ԣ���������ԡ������Զ������Ե���֯��ϸ������������֯������ϸ������֯��ϸ����λʱ�䣨ÿ���ӣ����ܹ��Զ������˷ܵĴ��������Զ��˷ܵ�Ƶ�ʣ��Ǻ����Զ������Ըߵ͵�ָ�ꡣ
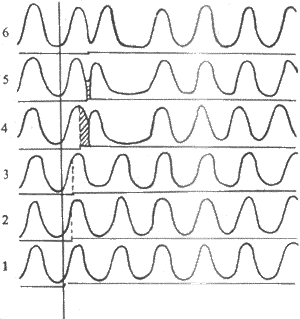
ͼ4-11 ��ǰ�����ʹ����Լ�Ъ
ÿ�������µĵ�ű�Ǻ���ʾ�����̼���ʱ�䣬����1-3��
�̼�������Ч��Ӧ���ڣ�������Ӧ������4-6��
�̼�������Բ�Ӧ���ڣ�������ǰ�����ʹ����Լ�Ъ
1���ļ����Զ������Ժ����ɼ���֯�����ϵ������ǰ�����˹۲쵽�������������£�������Ͳ����ද����������࣬��δ�ܵ��κδ̼�������£����Գ�ʱ��ء��Զ��ء��н���ؽ����˷ܺ����������ǣ�ֻ�е��˽���������ϸ�����缫������¼�Ŀ�Ĥ��λ�Ƿ����4���Զ�ȥ������һ��������ȷ�е�֤���������������ļ�ϸ������ֻ�������������֯��ijЩ����ϸ���ž����Զ������ԡ������ϵͳ������λ���������⣩���������еȼ����������ϸ����������ߣ��Զ��˷�Ƶ��ԼΪÿ����100�Σ�ĩ���ֿ�Ұ��ά����������ͣ�Լÿ����25�Σ��������ҽ��磨Լÿ����50�Σ��ͷ�����֧�����������ν�������֮�䡣
��һ������������������������м�����Ҫ���������塣��ô��������������֯�Ļ��ô��ͳһ�����������ڡ�����Ϊ�����أ�ʵ���к����۲쵽������ʼ�������յ�ʱ�������������ߵIJ�λ���������˷������л�ġ������˵�������ֵĻͳһ����������߲�λ����������֮�¡���������£������������ߣ����Զ��������˷��������������μ����ķ��������ҽ��硢�������������ڴ�����֯�����Ҽ����������������˷ܺ��������ɼ��������������������˷ܺ�������������λ���ʳ�Ϊ�����㡣������λ������֯�������ֳ������������Զ������ԣ�ֻ�������˷ܴ������ã��ʳ�ΪDZ���㡣��ij���쳣����£��������������֯�����磬���ǵ����������ߣ���������˷������Ͷ����ܿ���ijЩ������֯��Ҳ�����Զ������˷ܣ����ķ������������ӵ�ʱ����½�������߲�λ���˷ܶ���������Щ�쳣����λ���Ϊ��λ�㡣
�����DZ����Ŀ��ƣ�ͨ�����ַ�ʽʵ�֣�������ռ�졣��������Ը�������DZ���㣬���ԣ���DZ����4���Զ�ȥ����δ�ﵽ�е�λˮƽ֮ǰ�������Ѿ��ܵ��ᷢ�������δ����������˷ܵļ������ö������˶�����λ�����������Զ��˷ܾͲ����ܳ��֣��ڳ���ѹ�ֻ�������ѹ�֣�overdrive suppression���������DZ���㣬���ɲ���һ��ֱ�ӵ��������á����磬���������DZ����Ŀ���ͻȻ�жϺ����Ȼ����һ��ʱ�������ͣ����Ȼ�����Ҳ��ܰ�������DZ����Ľ��ɷ����˷ܺͲ�����������������ԭ���ǣ��������Ժܸߵ�����˷������£�DZ���㡰�������˷ܵ�Ƶ��ԶԶ�������DZ������Զ��˷�Ƶ�ʡ�DZ����ʱ��ġ����١��˷ܵĽ��������������ЧӦ��һ����������жϣ�����DZ������Ҫһ����ʱ����ܴӱ�ѹ��״̬�лָ��������������������Զ��˷ܡ�������Կ���������ѹ�ֵij̶����������Զ��˷�Ƶ�ʵIJ���ƽ�й�ϵ��Ƶ�ʲ����������ЧӦ��ǿ�������жϺ�ͣ����ʱ��Ҳ��������ˣ������˷�ֹͣ������������ɷ��ҽ����������Ϊ�㣬�����������Ҵ�����֯���ȴ��棻��Ϊ��ͷ��ҽ�����Զ��˷�Ƶ�ʲ���С������ѹ�ֵij̶Ƚ�С������ѹ�ֲ����Ļ��ƱȽϸ��ӣ�Ŀǰ��δ��ȫŪ�壻����һ��ʵ��ʾ���ǣ����˹�������£��������Ҫ��ʱ�ж�����ʱ�����ж�֮ǰ������Ƶ��Ӧ���������Ա��ⷢ���IJ���ͣ��
2��������Ӱ�������Ե���������ϸ�����Զ��˷ܣ���4��Ĥ�Զ�ȥ����ʹĤ��λ�������λ�ﵽ�е�λˮƽ������ġ���ˣ������Եĸߵͣ����������λ���е�λ�IJ���Ӱ�죬Ҳȡ����4��Ĥ�Զ�ȥ�����ٶȣ�ͼ4-12����
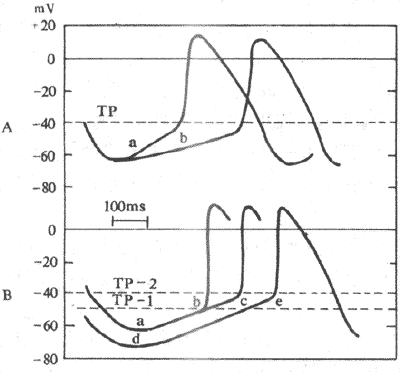
ͼ4-12 Ӱ�������Ե�����
A����λб����a���ٵ�bʱ�������Խ���
B�������λˮƽ��a�ﵽd�����е�λ��TP-1����TP-2ʱ�������Ծ����� Tp ���е�λ
��1�������λ���е�λ֮��IJ�ࣺ�����λ����ֵ���ٺͣ����е�λ���ƣ���ʹ����֮��IJ����٣��Զ�ȥ�����ﵽ�е�λˮƽ����ʱ�����̣����������ߣ���֮��Ȼ�����磬������ϵͳ�˷�ʱ��ʹ������ϸ��K+ͨ�����������ߣ����临��3����K+�������ӣ������λ����ֵ���������Խ��ͣ����ʼ�����
��2��4���Զ������ٶȣ�4���Զ������ٶ���Ĥ��λ�������λˮƽ�ﵽ�е���ˮƽ����ʱ��������أ��������ٶ����죬���е�λˮƽ����ʱ�����̣���λʱ���ڷ����˷ܵĴ������࣬���������ߡ���ǰһ����֪��4���Զ������ٶ�ȡ���ھ���������������ٶȣ���ȡ����Ĥ�ھ�����������ٶȡ����磬�����Ӱ�������ǿIf����������ֿ�Ұϸ��4�ڳ����ٶȣ�����������ԡ�
�������ļ��Ĵ����Ժ��������˷ܵĴ���
�ļ��ڹ�������һ�ֺϰ��壬�ļ�ϸ��Ĥ���κβ�λ�������˷ܲ�������������ϸ��Ĥ���������ҿ���ͨ�����̴��ݵ���һ���ļ�ϸ�����Ӷ����������ļ����˷ܺ�������������λ��ϸ��Ĥ�������ٶȿ���Ϊ���������Ե�ָ�ꡣ
1���������˷ܴ�����;�����ص� ����������ᷢ�����˷�ͨ���ķ����������������ķ������ķ��������������ķ�����ɵġ����ƴ���ͨ·��Ѹ�ٴ������ҽ�����������������������֧�����ֿ�Ұ��ά�����������Ҽ��˷ܣ���ֱ��ͨ�����Ҽ����˷�����Ĥ������Ĥ�����Ҽ��������������������˷ܡ����ڸ����ļ�ϸ���Ĵ����ԸߵͲ��ȣ��˷�������������ִ������ٶ��Dz���ͬ�ġ����ķ���һ���ķ����Ĵ����ٶȽ�����ԼΪ0.4m/s�����������ƴ���ͨ·���Ĵ����ٶȽϿ죬����˷ܿ���������Щͨ·�ܿ촫�������ҽ������������ң����Ҽ��Ĵ����ٶ�ԼΪ1m/s���������ڴ�����֯�Ĵ�����ȴ�ߵö࣬ĩ���ֿ�Ұ��ά�����ٶȿɴ�4m/s������������״�ֲ������ұڣ��������ɷ��ҽ��紫�����ҵ��˷ܾ����Ÿ��ٴ������ֿ�Ұ��ά��Ѹ�ٶ��㷺���������������ұڴ����������ԣ����ֶλ�Ŀ��ٴ������ڱ������ҵ�ͬ��������ʮ����Ҫ�ġ����ҽ�����ϸ���Ĵ����Ժܵͣ��������Խ�����ͣ������ٶȽ�0.02m/s�����ҽ���������ʱ�˷����ķ��������ҵ�Ψһͨ�������������ֻ�������ʹ�˷��������Ӹ�һ��ʱ�䣨�Ʒ�-���Ӹ飩�������Ҵ������Ӷ�����ʹ�������ķ��������֮��ſ�ʼ�����������ڲ������������ص��������Կ������������˷ܴ���;�����ص�ʹ����ٶȵIJ�һ���ԣ���������������д���ء�Э���ؽ��������������ʮ����Ҫ�����塣
2��������Ӱ�촫���Ե����� �ļ��Ĵ�����ȡ�����ļ�ϸ��ijЩ�ṹ�ص�͵��������ԡ�
��1���ṹ���أ�ϸ��ֱ����ϸ���ڵ���ʷ����ϵ��ֱ��С��ϸ���ڵ�������ľֲ�����С�ڴִ��ϸ�����˷ܴ����ٶ�Ҳ�Ϻ��������ķ��������Ҽ����ֿ�Ұϸ����ֱ��������ͷ��ҽ���ϸ�������У�ĩ���ֿ�Ұϸ����ֱ�������ijЩ���ֱ���ɴ�70��m�����˷ܴ����ٶ���죻��ϸ��ֱ����С��Լ5-10��m���������ٶȺ�����������ϸ��ֱ����С�������ٶ�Ҳ������
�ڻ������������У��ļ�ϸ��ֱ������ͻȻ�������Եı仯����ˣ���ֻ�Ǿ��������Ե�һ���ȽϹ̶������أ����ڸ���������ijЩ����������ļ������Եı仯��������Ҫ���á�
��2���������أ��ļ�ϸ���ĵ����������Ǿ�����Ӱ���ļ������Ե���Ҫ���ء����������˷�ϸ����ͬ���ļ�ϸ���˷ܵĴ���Ҳ��ͨ���γɾֲ�������ʵ�ֵģ��ο��ڶ��£�����ˣ����ԴӾֲ��������γɺ��ڽ�δ�˷ܲ�λĤ���˷�����������������Ӱ�촫���Ե����ء�������������������������ģ�Ϊ�˷���ŷֱ�������
������λ0�ڳ������ٶȺͷ��Ⱦֲ��������˷ܲ�λĤ0��ȥ��������ģ�0��ȥ�����ٶ����죬�ֲ��������γ�Ҳ�����죬�ܿ�ʹ�ʹ�ڽ�δ�˷ܲ�λĤȥ���ﵽ�е�λˮƽ�����˷ܴ������졣��һ���棬0��ȥ�����������˷ܺ�δ�˷ܲ�λ֮��ĵ�λ�������γɵľֲ�������ǿ���˷ܴ���Ҳ���졣�����ǣ�Ϊʲô�ֲ�������ǿ����Ӱ�촫���ٶȣ�������ǿ�ľֲ����������ľ������ʹ���˷ܲ�λ��Զ�����β�λ�ܵ��ֲ������Ĵ̼����˷ܣ����˷ܵĴ����Ͽ졣����ϸ��ֱ���������֮�⣬�ֿ�Ұ��ά�ȿ췴Ӧϸ��0��ȥ���ٶȺͷ������Ը����������Ӧϸ������ǰ�ߴ����ԱȺ���Ϊ�ߵ���Ҫԭ��
��֪�������ļ�ϸ��0��ȥ���ٶȺͷ��ȵIJ����Ҫ��Ĥ�ϣ�0�ڣ�����ͨ���Ĺ������ʾ�������ô��ͬһ�ļ�ϸ�����Կ췴Ӧϸ��Ϊ����0��ȥ���ٶȺͷ�������ʲô���ص�Ӱ�죿�������˷���ʱ�Ѿ�ָ������Na+ͨ������״�������ʧ�����״̬�Ǿ����˷���������ȱʧ�͵��µ���Ҫ���أ�Ҳ����˵�����˷��Զ��ԣ�Na+ͨ������״������ͨ���ܷ���ţ��˷��Ե����ޣ��Լ���������׳̶ȣ��˷��Եĸߵͣ����������һ�����ۣ�Na+ͨ������״��������Ĥȥ�����е�λˮƽ��ͨ�����ŵ��ٶȺ��������Ӷ�����Ĥ0��ȥ�����ٶȺͷ��ȡ�Na+ͨ�������ٶȺ�����������״����ΪNa+ͨ����Ч�ʻ�������ʣ�ͨ�����������ƿ��Ÿ��ʣ���ʵ��֤����Na+ͨ����Ч��Ҳ�ǵ�ѹ�����Եģ������������ܴ̼�ǰ��Ĥ��Ϣ��λֵ�������ط���Na+ͨ����Ч�ʣ���0��ȥ����������ʷ�ӳNa+ͨ�����ŵ��ٶȣ��뾲ϢĤ��λֵ�ĺ�����ϵ������ΪĤ��Ӧ���ߣ�ͼ4-13����Ĥ��Ӧ���߳�S�Ρ�������Ϣ��λֵ��-90mV������£�Ĥ�ܴ̼�ȥ�����е�λˮƽ��Na+ͨ�����ٿ��ţ�0��ȥ������ٶȿɴ�500V/s����Ĥ��Ϣ��λֵ������ֵ�����ͣ�ȥ������ٶ��½�����Ĥ��Ϣ��λֵ������ֵ����һ�����͵�Ĥ��Ϊ-60��-55mVʱ��ȥ���ٶȼ���Ϊ0����Na+ͨ����ʧ������ܿ��š��������������ΪNa+ͨ��Ч�ʵĵ�ѹ�������½�����Ҫ����ע����ǣ��ھ�ϢĤ��λֵ������ֵ���ܵͣ�Ĥ��-60��-55mV��״���£����Ĥ�ܵ��̼��������Ǹ�����������λ�仯�����Dz���һ��0��ȥ���ٶȺͷ��ȶ���С�Ķ�����λ��������Ϊ������������¿�Na+ͨ���Ѿ�ʧ�����Ca2+ͨ��δ��Ӱ�죬��ˣ�ԭ���Ŀ췴Ӧϸ����ʱ��������Ca2+�������µ�����Ӧ��λ��Ե�ʣ��˷ܴ����ٶ�Ҳ�����Լ��������������Ѿ���Ĥ0��ȥ��������ͨ�������˸�������������Na+ͨ��Ч�ʵ����䷶�롣
ͼ4-13 Ĥ��Ӧ����
���˾�ϢĤ��λ֮�⣬Na+ͨ�����ŵ��ٶȻ����ļ�ϸ�������������ʵ�Ӱ�졣���磬����Ӣ�ƿ�ʹĤ��Ӧ����������λ�����ᶡʹ֮������λ�������������Щҩ�������£�Na+ͨ������Ч����Ȼ�ǵ�ѹ�����Եģ����ǣ�ͬһ��ϢĤ��λˮƽ��0��ȥ������ٶȵ���ֵ������ͬ��ǰ�߸������������ߵ���������
Ĥ��Ӧ����ֻ�����˾�ϢĤ��λֵ��Na+ͨ�������ٶȼ�0��ȥ���ٶȵ�Ӱ�죬ʵ���ϣ���Na+ͨ������������������0��ȥ������Ҳͬ�������ھ�ϢĤ��λֵ��������ϢĤ��λ����£�Na+ͨ�����������ٶȿ죬���ҿ�������Ҳ�࣬������λ0��ȥ�����ٶȿ죬����Ҳ�ߣ�ͼ4-14��������ϢĤ��λֵ������ֵ�����£��������֧���������ȵ͵Ķ�����λ��ͼ4-14�ң���
ͼ4-14 ��ϢĤ��λ�Զ�����λ��֧�ٶȺͷ��ȵ�Ӱ��
S������̼�
�ڽ�δ�˷ܲ�λĤ���˷��� �˷ܵĴ�����ϸ��Ĥ�����˷ܵĹ��̣���ˣ�Ĥ���˷��Ա�ȻӰ���˷ܵĴ�����ǰ�������پ�ϢĤ��λ���������λ�����е�λ�IJ�༰���ڽ�δ�˷ܲ�λĤ�Ͼ���0��ȥ��������ͨ������״���Ǿ����˷��ԴӶ�Ҳ��Ӱ�촫���Ե���Ҫ���ء����������ʱ���˷��Խ��ͣ�����̼���ֵ���ߣ���ͬʱ��Ĥȥ�����е�λˮƽ����ʱ���ӳ��������ٶ���˼����������ڽ���λ�γɶ���̼�������ǰ�˷ܵ���������˷ܲ�λ�γɵľֲ������̼��ͽ�����ǰ�˷ܸ������֮ǰ�����ڽ���λ����������ǰ�˷ܵ���Ч��Ӧ���ڣ����������˷ܣ����´������ͣ���������ǰ�˷ܵ���Բ�Ӧ�ڻ����ڣ���������֧����������С�Ķ�����λ���˷ܴ�����֮�������ɼ�����Ӧ�ڵĴ��ڣ��ǿ��ܵ����˷ܴ����ϰ�����Ҫ���ء�
�����������ļ���������������ܵ�Ӱ��
֧�����������������ʶ��ļ���������������ܾ���������Ӱ�죬���Ƕ��ļ�������͵��������Ե�Ӱ�죬��Ҫ��ͨ����������ͨ���Ŀ��Ŷ�ʵ�ֵģ������ļ��������ܵĵ��ڻ�����Ƚϸ��ӣ��������ܵĸı䣬�������������������ı�ļ̷�ЧӦ֮�⣬������ͨ���������ƶ��������ܲ���ֱ��Ӱ�졣
��һ���������������������
�������˷�ʱ���ں���ά�ͷŵ���������������ļ�ϸ��Ĥ��M�͵��������壬��������ѹ�������Ա�ʱ���Դ����Ե�ЧӦ���о�֤���������������ձ����K+ͨ���Ŀ��Ÿ��ʣ��ٽ�����K+�������������ļ�ЧӦ����Ҫ���ơ�
K+�������ձ����ӽ�Ӱ���ļ�ϸ��������Ķ�����ڣ��پ�Ϣ״̬��K+���������ӽ����¾�Ϣ��λ����ֵ������ˣ���Ϣ��λ���е�λ�IJ�������ļ��˷��������½���������ϸ��������������K+�������ӵĽ���������λ����ֵ������һ���棬��4��K+���������ӽ�ʹIk˥�����̼������Զ������ٶȼ����������������ؾ������������Խ��ͣ���������������۸���������K+�������ӵ��¸������٣�������λʱ�����̣���Ч��Ӧ����Ӧ���̣����ڶ�����λʱ�����̣�ÿһ������λ�ڼ����ϸ����Ca2+����Ӧ���٣�����֮�⣬�����о������֣�����������ֱ������Ca2+ͨ������������Ca2+�������á����ڽ���ϸ����Ca2+�����٣��ļ�����������Ӧ���ͣ����ֳ����Ա���ЧӦ�����⣬������������˷�ʱ�����ҽ�������Ӧϸ��������λ���ȼ�С���˷ܴ����ٶȼ�������Ҳ��������������Ca2+ͨ��������Ca2+�����Ľ����
�������Ľ����Ͷ���Ӱ�������
�Ľ�����ĩ���ͷŵĵ�����ȥ���������أ������ļ�ϸ��Ĥ���������������������ϣ�ͨ��ij�ֻ��ƣ�ʹͨ�����������ữ���ı�Ĥ������ͨ���Ŀ��Ÿ��ʺ�������ϸ���ṹ�Ĺ��ܣ��������Ա��������Ա�ʱ�����Ա䴫����ЧӦ�����������ʷ��ڵ�ȥ���������غ��������أ��Լ���Դ�Ԧ����弤����Ҳ���������á����������ļ�ЧӦ�Ļ�����ԱȽϵ�һ������Ҫ��ͨ���ձ����Ĥ��K+ͨ���Ŀ��Ÿ��ʶ��������õģ������Ӱ������ñȽϸ��ӣ�����������������Ca2+ͨ���Ŀ��Ÿ�����һ��Ҫ����֮�⣬��ͨ����������ϸ������ϸ�����Ƶ����ļ��ĵ��������Ժ��������ܡ���������ú����û��ƹ������£��������Ӱ���ȥ���������ء��������أ��ܼ�ǿ����ϸ��4�ڵĿ�Ĥ�������Ifʹ4���Զ������ٶȼӿ죬���������ߣ���������Ӧϸ��������0��Ca2+������ǿ���٣��䶯����λ�����ٶȺͷ��Ⱦ����ӣ����ҽ������˷ܴ����ٶȼӿ죻�������Ӱ���ʹ������K+�������죬�Ӷ�ʹ�������̼ӿ죬������������̣���Ӧ����Ӧ���̡���Ӧ��������ζ��0������ͨ��������̼ӿ죬���������Ӱ�ʹ���˷ܷ���Ƶ�����ӵ����û���Э����ʹ���ʵ������ӣ��������Ӱ�ͨ����ǿ�ļ�����������ǿ�����ļ���������Ҳ�����ļ������ţ������û��ƱȽϸ��ӡ������ԣ������Ӱ����Ĥ�ͼ�����Ca2+ͨ�����Ÿ��ʵ����Խ�����ϸ����Ca2+Ũ�����ߣ��ṩ�˴�ʹ�ļ�����������ǿ����������һ���棬�����Ӱ��ִ�ʹ���Ƶ���Ca2+�����½����Ӷ������ļ��������������������Ӱ���ǰһ������ǿ�ں��ߣ���ˣ����������ļ������������������õķ����෴�����ֳ�������Ч������ǿ���������Ա������á��������Ӱ������£��ļ������ٶ����죬�������Ź������Լ�ǿ�����������Ӱ��ܾ��������Ե�һ��ЧӦ������ЧӦ�����������õĽ�������ȣ������Ӱ���ʹ���Ƶ���Ca2+���ͷ��������ӣ�������Ϊ�����Ӱ������˼��Ƶ���Ca2+������Ca2+���Ժܿ���뼡�Ƶ���ϵ�״̬�½����������Σ������Ӱ����������������ȡCa2+���ٶȣ����ɴ̼�Na+-Ca2+������ʹ��������ϸ���ų�Ca2+���١��ۺ��������������Ӱ������£������ڰ���Ca2+Ũ���½��ٶ����죬���Ƶ����ֺܿ���Ca2+���룬��Щ���ö������ڼ�ԭ��ά�д֡�ϸ��˿����⣬�ļ�����ԽϿ���ٶ�������Ź��̡�
�Ľ����������Ӱ����ļ��������Ƕ��ģ��ܸ��ӵģ�����ЩЧӦ�ǻ���ͳһЭ���ġ����Ľ������˷��������ʼӿ죬�Ӷ�ʹ�����ں������ڶ����̵�����£��ļ����ż��ٿ����ֲ����������������̴��������ҳ�ӯ����ĺ������һ���棬��������������ǿ���ļ�������ǿ�����죬��ˣ�������Ѫ��Ҳ�������������ڵ����̶����٣��ټ����˷ܴ������٣��ķ����Լ����Ҽ�������ͬ����������ǿ��Ҳ��ʹ�ķ������ҵ�����ǿ�����ӣ�������ά��������������������Ľ������˷�ʱ�������ʼӿ��ͬʱ�����������ӻ䣬�������������Դ�����ӡ�
�ġ�����ĵ�ͼ
���������壬���ᷢ����һ���˷ܣ���һ����;���ͽ��̣����δ����ķ������ң���������������˷ܣ���ˣ�ÿһ���Ķ������У�����������˷ܹ����г��ֵĵ�仯��������;���������ʱ��ȶ���һ���Ĺ��ɡ����������仯ͨ��������Χ�ĵ�����֯����Һ����ӳ��������棬ʹ�������λ��ÿһ�Ķ�������Ҳ�������й��ɵĵ�仯���������缫��������������һ����λ��¼�����������仯���ߣ������ٴ��ϼ�¼���ĵ�ͼ��electrocardiogram,ECG�����ĵ�ͼ��ӳ�����˷ܵIJ����������ͻָ������е������仯����������Ļ�е�������ֱ�ӹ�ϵ��
�ļ�ϸ���������仯���ĵ�ͼ����Դ�����ǣ��ĵ�ͼ�����뵥���ļ�ϸ���������仯���������Ե�����ͼ4-15������������������Ҫԭ�������¼��㣺�ٵ����ļ�ϸ����仯����ϸ���ڵ缫��¼���õ��ģ���һ�������缫����ϸ����������һ���缫���뵽ϸ��Ĥ�ڣ�����ĵ�仯��ͬһϸ����Ĥ����ĵ�λ��������ɲ��Ĥ�Ķ�����λ��Ҳ�ɲ��Ĥ�ľ�Ϣ��λ���ĵ�ͼ�ļ�¼����ԭ��������ϸ�����¼������ֻ�ܲ�����˷ܲ�λ���д����˷�״̬�IJ�λ֮��ĵ�λ��ھ�Ϣ״̬�£����Ǽ�Ĥ����λ�������˷�״̬��ʱ��Ĥ�����λ֮��û�е�λ�ϸ�����¼���߶����ʵȵ�λ�ߣ����ܼ������𣻢��ļ�ϸ����仯�����ǵ����ļ�ϸ���ھ�Ϣʱ���˷�ʱĤ�����λ�仯���ߣ����ĵ�ͼ��ӳ����һ���Ķ���������������������仯����ˣ��ĵ�ͼ��ÿһ˲��ĵ�λ��ֵ�����Ǻܶ��ļ�ϸ�������ۺ�ЧӦ������ķ�ӳ������ϸ���ڼ�¼����ͬ���ĵ�ͼ������������Ӽ�¼�������仯����ˣ��缫���õ�λ�ò�ͬ����¼���ĵ�ͼ����Ҳ����ͬ��
ͼ4-15�ļ�ϸ����仯�����볣���ĵ�ͼ�ıȽ�
A���ķ���ϸ����仯 V�����Ҽ�ϸ����仯
���������ĵ�ͼ�IJ��μ����������塡�ĵ�ͼ��¼ֽ���к��ߺ����������Ϳ���Ϊ1mm��С����¼�ĵ�ͼʱ�����ȵ��������Ŵ�����ʹ����1mV��ѹ�ź�ʱ������������ϲ���10mmƫ�ƣ�������������ÿһС���൱��0.1mV�ĵ�λ�����С���ʾʱ�䣬ÿһС���൱��0.04s������ֽ�ٶ�Ϊÿ��25mm������ˣ������ڼ�¼ֽ�ϲ������ĵ�ͼ�����ĵ�λ��ֵ�;�����ʱ�䡣
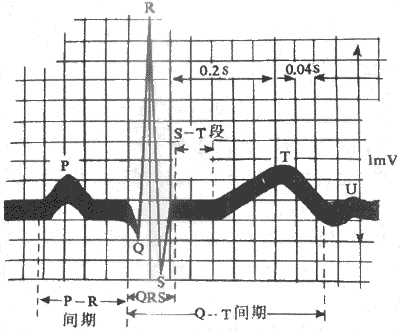
�����缫����λ�ú����߷�ʽ���Ƶ�����ʽ����ͬ����¼�����ĵ�ͼ���ڲ�����������ͬ���������϶�����һ��P����һ��QRS��Ⱥ��һ��T������ʱ��T��������һ��С��U����ͼ4-16����
ͼ4-16���������ĵ�ģʽͼ
1��P�� ��ӳ���������ķ���ȥ�������̡�P������С��Բ�ۣ���ʱ0.08-0.11s������������0.25mV��
2��Ta�����ķ�T���������ķ����������������ĵ�仯������ʼ��P��֮����P���ķ����෴��P-Ta���ڣ���P����ʼ��Ta�����˵�ʱ�̣�Ϊ0.15-0.45s����Ta����P-R�Ρ�QRS����ST�εij����ص���һ�𣬶���Ta�������ܵͣ���ͨ���ĵ�ͼ�Ͽ�����Ta����
3��QRS��Ⱥ��������������ȥ�������̵ĵ�λ�仯�����͵�QRS��Ⱥ�������������������ĵ�λ��������һ�����²�ΪQ�����Ժ��Ǹ߶����͵����ϵ�R���������һ�����µ�S�������ڲ�ͬ�����У�����������һ�������֡�����QRS��Ⱥ��ʱԼ0.06-0.10s���������Ҽ��˷����������ʱ�䣻���������ڲ�ͬ�����б仯�ϴ�
4��T�� ��ӳ���Ҹ��������Ҽ�ϸ��3�ڸ����������еĵ�λ�仯������һ��Ϊ0.1-0.8mV����R���ϸߵĵ�����T����Ӧ����R����1/10��T����ʱ0.05-0.25s��T���ķ�����QRS��Ⱥ������������ͬ��
5��U�� ��T����0.02-0.04s���ܳ��ֵ�һ���Ͷ����IJ�������һ����T��һ�£�����Լ0.1-0.3s�����������0.05mV���¡�U��������ͳ������ʮ�������
���ĵ�ͼ�У�����������������״���ض�������֮�⣬�����Լ�����֮���ʱ�̹�ϵҲ�������ۺ�ʵ�����塣���бȽ���Ҫ�������¼��
1��PR���ڣ���PQ���ڣ� ��ָ��P����㵽QRS�����֮���ʱ�̣�Ϊ0.12-0.20s��PR���ڴ�������������˷ܾ����ķ������ҽ���ͷ������������ң����������ҿ�ʼ�˷�����Ҫ��ʱ�䣬��Ҳ��Ϊ���Ҵ���ʱ�䣻�ڷ��Ҵ�������ʱ��PR�����ӳ���
2��PR�Ρ���P���յ㵽QRS�����֮������ߣ�ͨ�������ͬһˮƽ��PR���γɵ�ԭ���������˷ܳ嶯ͨ���ķ�֮���������Ҵ��������У�Ҫͨ�����ҽ��������˷�ͨ�����������dz��������γɵĵ�λ�仯Ҳ������һ���¼������������P��֮�������ֻص�����ˮƽ����ΪPR�Ρ�
3��QT���ڡ���QRS����㵽T���յ��ʱ�̣��������ҿ�ʼ�˷�ȥ������ȫ��������Ϣ״̬��ʱ�䡣
4��ST�Ρ�ָ��QRS��Ⱥ���˵�T�����֮��������ƽ����߶Σ����������Ҹ������ļ�ϸ�������ڶ�����λ��ƽ̨�ڣ�2�ڣ���������֮��û�е�λ����ڣ������ָֻ�������ˮƽ��