4 碳水化物
4.1 碳水化物的分类、结构和性质
4.1.1 碳水化物的分类
碳水化物分单糖、双糖、低聚糖、多糖四类。糖的结合物有糖脂、糖蛋白、蛋白多糖三类。
(1)单糖不能水 解成更简单的糖的碳水化物叫单糖。单糖有3到7个碳原子,依碳原子数的多少,依次称为丙、丁、戊、已、庚糖。丙糖和丁糖以中间代谢物的形式存在,自然界存在最多是戊糖和已糖。
单糖具有醛基或酮基。有醛基者称醛糖,有酮基者称酮糖。
(2)双糖每分子能水解成两分子单糖的碳水化称双糖。营养上有意义的双糖有蔗糖、麦芽糖、乳糖三种。
(3)低聚糖每分子水解成3~8个分子单糖的碳水化物称低聚糖,也有人把水解成3~10个,甚至20个分子单糖的碳水化物归入这一类。可再分两类:水解产生的所有糖分子都是葡萄糖的称麦芽糖低聚糖,由3个葡萄糖分子组成的叫麦芽三糖,四个葡萄糖分子组成的叫麦芽四糖等等。另一类水解时产生不止一种单糖,称杂低聚糖。如大豆中的杂低聚糖水解产生棉子糖和木苏糖等,人不易消化,无法利用。但机体自己合成的杂低聚糖,有很重要的生理功用。
(4)多糖每分子更多分子的单糖的碳水化物是多糖。可再分α葡聚糖、β-葡聚糖、杂多聚糖三类。α-和β-葡萄聚糖都是葡萄糖分子组成的,但联结的方式不同,前者是以α-糖苷键联结的,后者则用β-糖苷键。α-葡聚糖有淀粉、糊精、糖原三类。组成杂多聚糖的不止一种单糖,膳食纤维属这一类,除纤维素外,膳食纤维还包括半纤维素、果胶、藻类、木质素。
(5)糖脂糖和脂质的结合物称糖脂。简单可分两类:脑苷脂和神经节苷脂。
(6)糖蛋白和蛋白多糖广义讲,碳水化物和蛋白质结合后的化合物都是糖蛋白。糖蛋白的种类很多,并有广泛的生理功用。30年代以来,好几个科学家提出了不同的分类方法。
1946年Stacey把蛋白质含量低,以碳水化物的化学反应为主的称粘多糖;蛋白含量高,化学反应以蛋白质为主的称粘蛋白。
1962年Gottschalk根据碳水化合的结构分成两大类。简言之,有重复出现的双糖单位的低聚糖链或多糖链,与蛋白质结合者称多糖蛋白,后人把它叫做Protegolycans,这个名字一直沿用至今,译作蛋白多糖。以没有重复出现双糖单位的低聚糖单位的低聚糖链与蛋白质相结合的称糖蛋白。
80年代以来的分类,从化学结构出发,分得很细,不予介绍了。本章的讨论,以Gottschalk的分类法为主。
粘多糖和粘蛋白的名字,现在营养学书上仍有出现,但含义与Stacey提出的略有不同。粘多糖(Mucopolyscaccharides)指Gottschalk分类的蛋白多糖中的碳水化物部分,或称氨基多糖(Glycosaminoglycans)。粘蛋白指Gotrtschalk分类中的糖蛋白。有时把糖脂、糖蛋白和蛋白多糖一起,统称糖结合物(Glycoconjugates)。
4.1.2 碳水化物的化学组成和结构
(1)单糖单糖中的最重要的是葡萄糖,它的结构式有五种表示方法,见图4-1。
葡萄糖有六个碳原子,以阿拉伯数字计数如图。在图4-1(A)中可看出第2、3、4、5个碳原子所连的四个基都不相同,是不对称碳原子。每个不对称碳原子有两个异构体,羟基在左面或右面代表两个不同的化合物。习惯上把与伯醇基[图4-1(A)中第6个碳原子]相连的碳原子的羟基在右面的称D系的糖。图4-1(A)中用虚线框出的部分是D系糖必备的结构。所以,图4-1(A)代表的是D-葡萄糖。
图4-1(A)所示的开链式结构可以环合起来,即第5个原子上羟基的氢移到第1个碳原子的羟基氧上,然后连成环,见图4-1(B)。这样,第1个碳原子也变成不对称的的。羟基在右面的称α,D-葡萄糖,羟基在左面的称β,D-葡萄糖。
但是六个碳原子是不在一条直线上的,六个碳原子的环是六角形的。为了区别不对称碳原子的构型,可把这个环表示如图4-1(C)。环的平面垂直纸面,粗线表示在环平面前面的链,细线代表的链在环平面的后面。图4-1(B)中右侧的
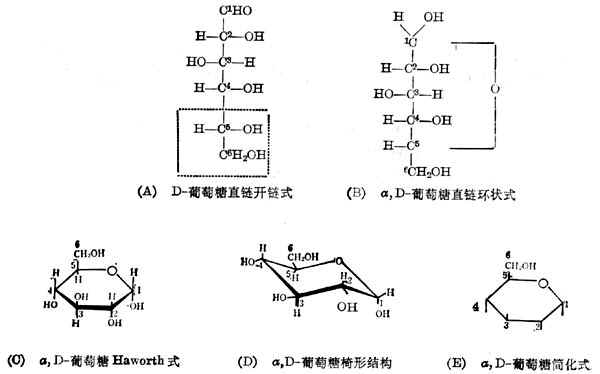
图4-1 葡萄糖的结构式
羟基写在环的平面之下,左侧的羟基写在环的平面之上。这样表示的结构式叫做Haworth投影式。
事实上,环的六个碳原子并不在一个平面上,X线衍射的结果说明它们是以椅形存在的,如图4-1(D)。但这样的表示方法书写困难,所以一般采用Haworth投影式。并且常简化如图4-1(E),这里把碳和氢都省了,仅用一条垂线表示羟基。现在国际上有一种通用的符号来代表糖及其衍生物,葡萄糖的是Glc。
自然界存在的葡萄糖,不论在单糖、双糖、低聚糖或多糖中,都以图4-1(D)的形式存在。只有在水溶液中,才有约1%以开链的形式存在。
糖有旋光性。在旋光计中,有的能使偏振光的平面右旋,有的使之左旋。命名时,右旋用(+)表示,左旋用(-)表示。不是所有的D系的糖都是右旋的。α,D-葡萄糖是右旋的,它的全名是α,D(+)葡萄糖。一般提到的葡萄糖,除特别指出的以外,都是α,D(+)葡萄糖。
果糖是D系的、β异构体、左旋的,全名是β,D(-)果糖。它的环由五个碳原子组成,在同一平面上。
图4-2 β,D(-)果糖
半乳糖是葡萄糖的异构体,所不同的是第4个碳原子上的羟基在环的上面,而葡萄糖的在环的下面。符号Gal。
(2)双糖在葡萄糖的环状结构中,第一个碳原子上有一个羟基。它可和另一个有羟基的化合物失水而成为糖苷,形成的键叫糖苷键。
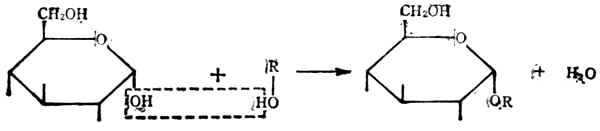
葡萄糖 葡萄糖苷
糖苷是自然界存在的一大类化合物,有些可作为药物。
如ROH是一个单糖分子,这样结合的就成为双糖。如两个单糖分子都是葡萄糖,以α-1,4糖甘键(即第一个葡萄糖分子的第1个碳原子上的羟基和第二个葡萄分子的第4个碳原子上的羟基脱水相结合而形成的键,亦写作α1→4)结合后得到的是麦芽糖。
蔗糖是α,D-葡萄糖和β,D-果糖以α-1→2糖苷键结合的。乳糖是β,D-半乳糖以β-1→4糖苷键和α,D-葡萄糖相结合的。
(3)低聚糖 麦芽三糖到麦芽八糖,都是α,D-葡萄糖以α-1→4和α-1→6糖苷键结合的。杂低聚糖匠结构比较复杂,将于糖蛋白中加以介绍。
(4)多糖
①淀粉淀粉是由许多葡萄糖分子组成的。与水调成糊状并加热到60~80℃后,可分成直链淀粉和支链淀粉两部分。前者约占15%,后者约占85%。
直链淀粉也由葡萄糖分子,以α-1→4苷键连成一条直链,并蜷曲如图4-3。
支链淀粉也由葡萄糖分子组成,它们不仅以α-1→4糖苷键连成直链,还有分支。分支处是以α-1→6糖苷键结合的。每一支有20~30个葡萄糖分子。图4-4是它的示意图。图4-5是图4-4中框出部分的放大,用以表示葡萄分子间的联结方式。
图4-3 直链淀粉示意图
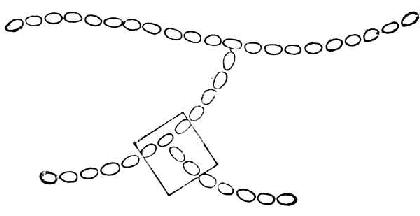
图4-4 支链淀粉示意图
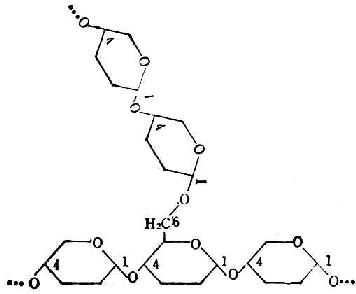
图4-5 葡萄糖分子在支链淀粉中的结合方式
②糖原糖原由6000~3000个葡萄糖分子组成,结构式和支链淀粉相似,不过分支更多。每一分支约有11~18个葡萄糖分子。分支处是α1→6糖苷键,其余是α1→4糖苷键。
③糊精和麦芽低聚糖二者都是淀粉水解的中间产物。结构中有α1→4和α1→6糖苷键。
④纤维素纤维素是β-葡萄糖以β1→4糖苷键连成直链,每链约有2800个葡萄糖分子。结构如图4-6,图中葡萄糖分子的环,是前、后交替地与纸的平面垂直的。
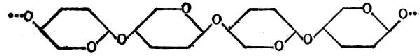
图4-6 纤维素的结构式
⑤膳食纤维
半纤维素是D-甘露糖、D-木糖、D-葡萄糖、D-半乳糖以β1→4糖苷糖连起来的,分支上有阿拉伯糖、半乳糖和葡萄糖醛酸。
果胶主链葡萄糖醛酸连成的,侧链上有鼠李糖、阿拉伯糖、岩藻糖、木糖。
藻类多糖主链由甘露糖、木糖、葡萄糖醛酸和葡萄糖组成,侧链有半乳糖。
木质素木质不是碳水化物,是苯丙烷的聚合物。由于和其余膳食纤维同时存在,且有类似的功用,所以把它作为膳食纤维中的一类。
膳食纤维和粗纤维不同。植物性食物中,有胃肠道不能消化的物质,统称膳食纤维;食物用酸碱处理后的不溶物称为粗纤维。经过这一处理,大部分膳食纤维都丢失,测得的数值仅有膳食纤维总量的20~50%。
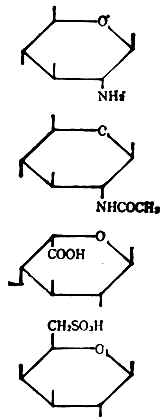
⑥单糖的衍生物低聚糖和多糖不仅能由单糖组成,还能由单糖的衍生物所组成。表4-1列出了它们的类别,每类举一例表示结构式及其名称和符号。
表4-1 单糖的衍生物
| 类别 | 结构式 | 名称 | 符号 |
| 氨基糖 |
| β-D氨基葡萄糖 | ΒDGlen |
| 乙酰氨基糖 | N-乙酰氨基-β-D葡萄 | ΒDGlcNAc | |
| 糖醛酸 | α-L-艾杜糖醛酸 | Αltdu | |
| 硫酸酯 | 4-硫酸-β-D-半乳糖 | ΒDGal-6SO3h |
另有一类组成低聚糖和多糖的单糖衍生物,叫做唾液酸,除有乙酰氨基和羟基外,还和甘油连在一起。结构式见图4-7,符号NeuAc。
糖的醛基被氧化后,得糖酸如D-葡萄酸(图4-8)。醛基被还原成羟基后,得糖醇如山梨醇(图4-9)。
⑦糖脂糖脂的基本结构是鞘氨醇(图4-10),它的氨基与脂肪酸(含14~16个碳原子)酰化后得脂酰鞘氨醇,符号为Cer。再与葡萄糖或半乳糖相连成脑苷脂。和低聚糖(由几个或多至30多个单糖或其衍生物组成)相连后成神经节苷脂。已确定结构式的有数十种,区别在于糖链和脂肪酸的不同。
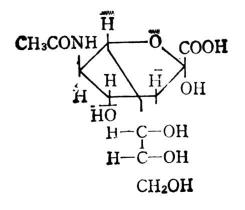 | 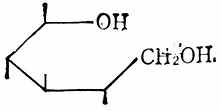 | 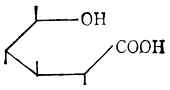 |
| 图4-7 唾液酸 | 图4-8 D-葡萄酸 | 图4-9 山梨醇 |
糖脂的糖链,和蛋白多糖的糖链常有一部分相同。所以它们的生理功用,有不少相似之处。
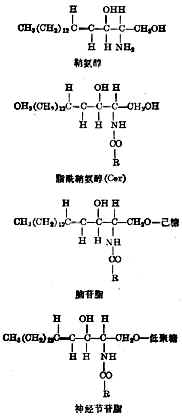
图4-10糖脂的化学结构
⑧糖蛋白中低聚糖糖蛋白中的单糖或基衍生物有以下种:
戊糖:阿拉伯糖(Ara)、木糖(Xy1);
甲基戊糖:岩藻糖(Fuc);
已糖:葡萄糖、半乳糖、甘露糖(Man);
乙酰氨基已糖:乙酰氨基葡糖(GlcNAc)、乙酰氨基半乳糖(GallNAc);
唾液酸。
不同的糖蛋白,由这些单糖或其衍生物中的一部分,组成不超过15个单体的低聚糖链,再和蛋白质结合而成。糖含量为1~85%。
⑨蛋白多糖中的聚氨基糖这一类糖是多糖。糖链中有交替出现的糖醛酸(葡萄醛酸或艾杜糖醛酸)和氨基已糖(氨基葡萄糖或氨基半乳糖或基乙酰化产物)。习惯上说成有交替出现的双糖单位。共分七类,见表4-2。
在这七类中,透明质酸有羧基,其它六类都有硫酸基,因此聚氨基糖都是多价的阴离子。
究竟有多少单糖分子聚合成聚氨基糖?根据来源而不同,分子量自4000到1000000以上。
表4-2 蛋白多糖中的氨基糖的分类和组成
| 聚氨基糖 | 双糖单位 | 取代基 | 其它单糖 | |
| 糖醛酸 | 氨基已糖 | |||
| 透明质酸 | βDGel | βDGleNAc | Gal;Xyl | |
| 4-硫酸软骨素 | ΒDGel | ΒDGalNAc | GalNAc 6位硫酸化 | Gal;Xyl |
| 6-硫酸软骨素 | ΒDGel | βDGalNAc | GalNAc 6位硫酸化 | |
| 硫酸皮肤素 | βDGal αLIeU | αDGlcN | 部分IdU2位硫酸化 部分GalNAc4或6位硫酸化; | Gal;Xyl |
| 硫酸类肝素 | ΒDGal αLIdU | αDGlcN | GlcN氨基乙酰化或硫酸化; 部分GlcN6位硫酸化; 部分IdU2位硫酸化 | |
| 肝素 | βDGal `αLIdU | αDGlcN | GlcN氨基乙酰化或硫酸化; 部分GlcN3或6位硫酸化; 部分IdU2位硫酸化 | Gal;Xyl |
| 硫酸角质素 | βDGal | βDGlcNAc | 部分GleNAc6 | GalN;Man; Fuc;NeuAc |
透明质酸单独存在,一般都是单独的一条多糖链。其它六类中,除硫酸角质素外,都通过半乳糖-木糖和蛋白质的丝氨酸相连结的,一条肽链上可连几条糖链。
4.1.3 碳水化物的物理性质
(1)溶解度单糖、双糖、低聚糖、糊精都溶于水。淀粉不溶,与水加热后可吸水膨胀,变成糊状。淀粉经酸处理生成可溶淀粉。糖原能分散在水中得乳白色胶态“溶液”。纤维素不溶。果胶能溶于水。其余膳食纤维可吸水膨胀,吸水量依来源、周围液体的PH和离子浓度等而不同。如麦麸可吸收5倍生本身重量的水。吸水后膳食纤维呈海绵状,细菌和一些分子能穿插进去。糖脂糖的一端亲水,脂质的一端疏水。糖蛋白和蛋白多糖的糖也都亲水。
(2)甜度如以蔗糖的甜度作100,一些糖及糖醇的甜度见表4-3。其中转化糖是蔗糖的水解产物,含葡萄糖和果糖各50%。
表4-3 糖的甜度
| 糖或其衍生物 | 甜度 |
| 果糖 | 173 |
| 转化糖 | 130 |
| 蔗糖 | 100 |
| 葡萄糖 | 74 |
| 山梨醇 | 60 |
| 甘露醇 | 50 |
| 半乳糖 | 32 |
| 麦芽糖 | 32 |
| 乳糖 | 16 |
4.2 碳水化物的生化代谢
4.2.1 消化吸收
碳水化物要消化或单糖才吸收。消化的过程就是水解的过程。麦芽糖、乳糖、蔗糖、麦芽低聚糖都能消化。人能消化的多糖仅淀粉一种,糖原在制成食品时已不存在了。
消化从口腔开始,口腔里有唾液淀粉酶能水解交替α1→4糖甘键,但不能水解α1→6糖苷键和相邻的α1→6糖苷键。消化产物是糊精、麦芽低聚糖和麦芽糖。
胃里没有消化淀粉的酶。唾液淀粉酶的最适PH是6.6~6.8,在食糜没有被胃酸中和以前,能持续作用一段时间,使淀粉和低聚糖能再消化一部分。
小肠内有胰液的α-淀粉酶,其作用和唾液淀粉酶相同,把直链淀粉消化成麦芽糖和麦芽三糖,支链淀粉消化成麦芽糖、麦芽三糖及由4~9个葡萄糖分子组成的而有α1→6苷键的麦芽低聚糖。
肠粘膜上皮细胞中有吸收细胞,每一细胞约有3000条微绒毛,微绒毛间的空间的有效半径约0.4nm。只有上述消化产物能够通过,与微绒毛膜上的酶反应。膜上的酶有四种:①α1→4糖苷酶,把葡萄糖分子自上述产物一个个地切下来;②异麦芽糖酶,水解麦芽低聚糖的α1→6糖苷键;③蔗糖酶,消化蔗糖;④β-半糖苷酶,消化乳糖。
所以消化分两步进行:①肠腔内的消化,产物是双糖和麦芽低聚糖;②微绒毛膜上的消化,产物是单糖。
微绒毛双脂质层的(详后)。四种酶嵌在双脂质层内,活性位伸在膜外。在其近处,还有全部嵌在膜内的运输单糖的蛋白质,这样,消化的最终产物立刻可以被运输蛋白所结合。
运输蛋白在结合葡萄糖以前,先结合肠腔内的Na+排入肠腔,肠腔中Na+都带入细胞内,释放到胞浆中。Na+排入肠腔,肠腔中的Na+浓度比细胞内高,自低浓度排到高浓度要消耗能量,所需能量由ATP供应。糖进入细胞后,约有15%流回肠腔,25%扩散入血,60%与靠近基膜一端的质膜上的另一载体蛋结合而离开细胞。这一结合不需Na+,而且运输葡萄糖的速度比葡萄糖从肠腔进入吸收细胞的速度快,所以葡萄糖不会在吸收细胞中蓄积,从而提高了吸收效率。当食糜到达空肠下部时,95%的碳水化物都被吸收了。
4.2.2 中间代谢概述
小肠吸收的碳水化物主要是葡萄糖、果糖、半乳糖三种,经门静脉送到肝脏。葡萄糖进入肝细胞后与磷酸反应生成葡萄糖-6-磷酸,这样细胞内的葡萄糖浓度可维持在低水平,使葡萄糖不断进入肝细胞。在吸收的葡萄糖中,60%以上在肝内代谢,其余入大循环。果糖和半乳糖在肝中转变为葡萄糖。
葡萄糖在肝内经分解代谢提供机体所需要的能量,多余的合成糖原保留在肝内,再有多余就转变成脂肪运送到脂肪组织储存起来。
葡萄糖的分解代谢分三个阶段:
(1)酵解葡萄糖分解的第一阶段是生成丙酮酸。这一系列反应和碳水化物在体外经酵母的作用生成丙酮酸相同,所以称为酵解。所不同的是发酵时丙酮酸进一步生成洒精;在体内,有氧时丙酮酸氧化成二氧化碳和水,无氧时还原成乳酸。
酵解分十步进行(图4-11)。为了要把葡萄糖的6个碳原子一分为二,先使它两端磷酸化,才能断列成两个接近相等的部分,然后脱去磷酸成丙酮酸。
上述反应都在细胞浆中进行。每一步都有特定的酶参加。第①、③两步消耗2分子ATP,说明葡萄糖分解代谢开始时要用能量来推动。⑥、⑨两步各产生2分子ATP。从葡萄到丙酮酸,净生2分子ATP,酵解产生的能量不多。
细胞浆中NAD+和NADH的总量基本恒定。如所有的NAD+都变成NADH,酵解就得停止。要使葡萄继续不断氧化成酮酸,必需把NADH的氢转移到另外的受体上去。有氧时氢经呼吸链氧化成水。无氧时氢交给丙酮酸使成乳酸⑩。乳酸过多时离开细胞入血。所以严重缺氧者可发生代谢性酸中毒。
(2)丙酮氧化成乙酰酶a 有氧时,丙酮酸进入线粒体。在丙酮酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酶A,同时产生1molATP和1molNADH。维生素B1B2、烟酰胺、泛酸参加。反应是不可逆的。
(3)三羧酸循环乙酰辅酶A上的两个来自葡萄糖的碳原子,经三羟酸循环生成2molCO2,完成葡萄糖的氧化。反应在线粒体中进行。这一过程分9步(图4-12)。
三羧酸循环中产生3molNADH,1molFADH2,1molGTP。
NADH把氢通过FAD和细胞色素交给氧,同时放出能量。每molNADH通过这一系列反应所放出的能量是220kJ(52.6kCal)。这些能量以ATP的形式贮存起来。每molATP能产生30.5Kj(7.3kCal),理论上每molNADHRNUD氧化产生的能量可用来合成7molATP。这是指在标准状况(25℃,1个大气压,作用物和产物浓度都是1M/LPH7)下说的。在模拟体内的条件时,实验证明只能合成3molATP。如用反应式表示:
NADH+H++1/2O2

3ADP+3H3PO4+能量
前面一个是氧化反应,后面一个是磷酸化,二者偶合在一起,称为氧化磷酸化。
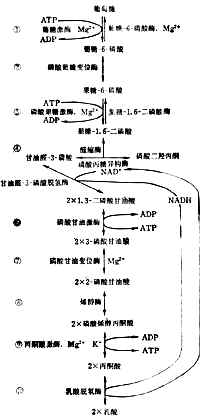
图4-11 糖的酵解
实验也证明FADH2氧化时产生2个ATP,GTP可生成1molATP。根据这些数值,葡萄糖完全氧化时,机体获得的能量可归纳如下:
①酵解:葡萄糖→2丙酮酸+8ATp
②丙酮酸氧化成乙酰辅酶A:
2丙酮酸→2乙酰辅酶A+2CO2+6ATp
③三羧酸循环:
2乙酰辅酶A→4CO2+24ATp
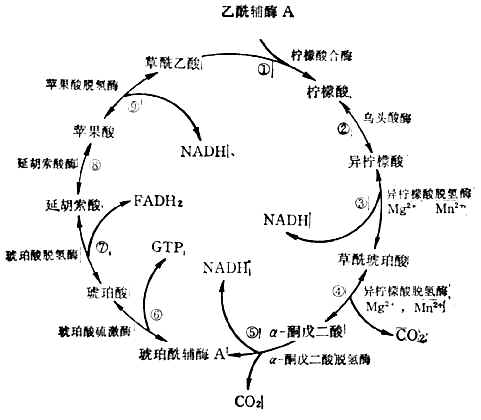
图4-12三羧酸循环
即每mol葡萄糖在体内氧化成6molCO2时可产生38molATP,机体可以利用的能量是38×30或1140kJ[(38×7.3)或277kCal]。每mol葡萄糖氧化成6molCO2和水时,自由能的改变是2870KJ(686kCal)。所以,机械效率=1140/2870×10=40%。
这是利用标准状况的数字计算的,在体内高一些,约60%。
从上面归纳的数字也可看出碳水化物提供的能量,在氧化的三个阶段中,以三羧酸循环最多。脂肪和蛋白质最后也通过三羧酸循环氧化。食物向机体提供的能量,90%来自三羧酸循环。
葡萄糖代谢另外还有一种途径,不通过果糖二磷酸,叫做已糖一磷酸通路。由于产生核榶,又叫做戊糖-磷酸通路。
(4)已糖-磷酸通路这一通路目的是提供合成核酸所需的核糖,以及合成脂肪酸、胆固醇等所需的能量供应者NADPH(还原型辅酶Ⅱ)(图4-13)。肝脏中约有30%的葡萄糖经过这一通过代谢。红细胞中这一通路也很活跃。它的失调可导致溶血性贫血。反应都在细胞浆中进行。
核糖-5-磷酸既可用于核酸的生物合成,又可与5-磷酸木酮糖反应生成甘油醛3-磷酸和果糖-6-磷酸。究竟走哪一条路,要看细胞本身对NADPH和核糖的需要量而定。许多细胞NADPH要得多,过剩的戊糖可经甘油醛-3-磷酸和果糖-6-磷酸酵解,或由果糖-6-磷酸转变成葡萄糖-6-磷酸而重走这一通路。
这一通路需维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1。5-磷酸核酮糖转变为葡萄糖-6-磷酸的反应,可用于评价维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1的营养状况。
(5)糖原葡萄糖到各个细胞发挥作用以后,多余的可储存起来。避免细胞内渗透压的升高,细胞把葡萄糖合成大分子的糖原。需要能量时糖原再分解葡萄糖。如肝糖原产生的葡萄糖,用以维持血糖的恒定。肌糖原产生的葡萄糖,直接用于供给能量。
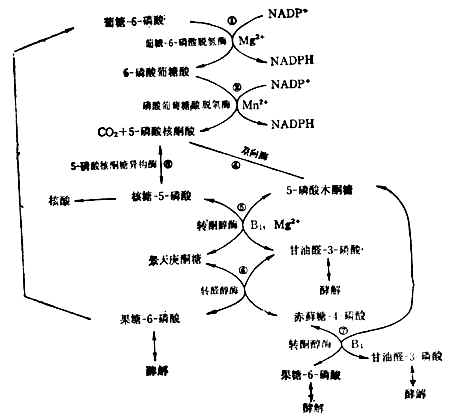
图4-13 已糖-磷酸通路
4.2.3 三大营养代谢的关系
人习惯于三餐,饱腹和空腹交替产生。所以要具备能处理碳水货物增多和缺少时保证能量供应的两套本领。
餐后血糖升沿高,胰岛素分泌增加,胰高血糖素分泌减少。使更多的葡萄糖进入肝脏、肌肉和脂肪组织;增强酶活力,加速葡萄的氧化和肝糖原、肌糖原的合成。超过糖原的储存量后,肝脏可把葡萄糖经磷酸二羟丙酮还原成甘油-3-磷酸,与乙酰辅酶A合成的脂肪酸,利用NADPH提供的能量,生成脂肪。再和蛋白质结合成极低密度脂蛋白入血,运送到脂肪组织储存。过多的碳水化物也能合成某些非必需氨基酸。
餐后约4h,不能利用食物中的葡萄糖。胰高血糖素分泌增加,胰岛素减少,转为利用肝糖原以供给血糖。肝糖原储存量不多,成人能动用的不到70g,不够一个晚上的消耗。所以,从餐后8h开始,还得利用脂肪以提供能量。甘油可变成糖,但在脂肪中所占重量不多。脂肪酸只能供能,不能转变为葡萄糖。因此,血糖的来源不得不转向氨基酸了。一般说来,除亮氨酸和赖氨酸外,其他氨基酸都是能成糖的,但从能量的观点看,只有丙氨酸和谷氨酸可作葡萄糖的来源。此时血糖的90%来自丙氨酸,10%来自谷氨酸。
肝脏能把肌肉中送来的丙氨酸作用成丙酮酸而生成葡萄糖。空腹时,肌肉中进入血液的氨基酸增加,其总量的30~40%是丙氨酸。但肌肉蛋白质的丙氨酸含量不到10%。另外的丙氨酸从哪里来的?葡萄糖经酵解可生丙酮酸,再经转氨作用丙氨酸。但这样葡萄糖的量并没有增加。增加的丙氨酸实际来自其他氨其酸。
肾脏把谷氨酸去氨后得α酮戊二酸,再和运来的成糖氨基酸如丙酸起转氨作用而得丙酮酸,最后得葡萄糖。α-酮戊二酸也能经三羟酸循环形成苹果酸,最后成葡萄糖。
4.3 碳水化物的生理功用
4.3.1 供给能量
(1)产热量每mol葡萄糖氧化供能时,理论上最多得2870Kj(686 kcal)。葡萄糖的分子量是180,每克葡萄糖产热2870/180=15.94kJ(686/180或3.81kcal)。传统上取其整数16Kj(4kcal)。世界卫生组织,把葡萄糖订为16KJ(3.75kcal),淀粉为18Kj(4.1kCal)。我国和美国药典规定配制输液用的葡萄糖含1mol结晶水,每克产热16×180/198或14.5kJ(3,75×180/198或3.41kCal)。静脉输液时葡萄糖提供的能量应以14.5Kj/g(3.41kCal/g)计算。
(2)膳食中碳水化物所占能量的百分比碳水化物没有规定的需要量,因为氨基酸和甘油在体内都能变成糖。但每人每天至少需摄食可以消化的碳水化物为50~100g。否则会引起酮病、组织蛋白分解过多,以及阳离子,特别是钠离子,和水的丢失。
食物中碳水化物太多也不好。根据营养学家的意见,总能量中的55%应由碳水化物来供应,其中单、双糖提供的不多于14%。
摄入多糖(主要是淀粉)的同时,能获得蛋白质、脂类、维生素、矿物质、膳食纤维。摄入单双糖(主要是蔗糖)时,不能取得除糖以外的其他营养素。而且摄入蔗糖过多能引起龋齿、心血管疾病和糖尿病。
(3)蔗糖过多的影响
①冠心病:蔗糖过多,膳食纤维过少,冠心病的死亡率高。如本世纪以来,美国人的死亡率越来越低,而冠心病死亡率却明显升高。自1900年的每10万人中167.3人,增至1965年的237。7人。
从1909年到1965年,美国人的食物中,每人每天摄取的能量下降约10%。蛋白质的变化不大。脂肪在总量中所占的百分数,虽增加9%,但增加的主要不不饱和脂肪酸(以亚油酸为代表)。从食物中计算所得的胆固醇摄入量,1909~1913年每人每天是495mg,1965年是518mg。所以脂肪和胆固醇变化不大。最明显的变化是淀粉摄入减少,而单、双糖吃得多了。1909年~1913年每人每到摄取碳水化物492g,其中蔗糖占31.7%;1965年碳水化物374g,蔗糖占51.2%。
再从膳食纤维的角度看,半个世纪以来,美国人的摄入量也有所减少。他们吃的水果和蔬菜的总量不变,水果的品种从苹果改为多吃柑桔。苹果含量胶多,柑桔的果胶集中在桔皮中,一般不吃。这一改变导致了膳食纤维摄入量的减少。
从横的方面,Yudkin比较了15个国家的能量、总脂肪、动物脂肪、植物脂肪、奶油、人造奶油、总蛋白、动物蛋白、蔗糖的摄取量和冠心病死亡率的关系,发现只和蔗糖成正相关。
纵横两个方面的比较,都说明摄取量和冠心病死亡率高,膳食纤维也有一定影响。不少实验研究也都证明这一点。
②糖尿病:Cohen把新移居以色列和长期定居在以色列的也门人的糖尿病发病率和碳水化物摄入量作了比较(表4-4)。说明蔗糖摄入多者,糖尿病发率高得多,而碳水化物总量和总能量在两组中出入不大。
表4-4 两组犹太人碳水第物摄入量和糖尿病发率的比较
| 组别 | 总能量(kCal) | 碳水化物总量 | 蔗糖 | 糖尿病发病率,% | ||
| Kcal | ||||||
| 移居10年以内者 | 2237 | 343 | 1372 | 6.6 | 26.4 | 0.06 |
| 移居25年以内者 | 2559 | 377 | 1508 | 63.0 | 252.0 | 2.90 |
蔗糖多吃不好,那么究竟应吃多少?对健康人,每天每公斤体重2g,短期内不会增加血糖、甘油三酯和脂肪酸。心脏病和糖尿病患者,蔗糖所提供的能量占总能量的5%以下无害。
(4)营养补给时碳水化物的选择口服碳水化物补充能量时,不能选单、双糖。如需供能8400Kj(2000kCal),用葡萄糖时需586g。但人对水的耐受量仅3L,因此必然要用近20%的溶液,这样它渗透压是血清的4倍,会引起腹胀、腹痛、腹泻。如选平均分子量是葡萄糖5倍的麦芽低聚糖,则25%时等渗,不会产生上述高渗性副作用。这种低聚糖很容易消化吸收。
周围静脉输液时,渗透压不能高于500mOsm·kg-1水,否则会引起血栓性静脉炎。10%葡萄糖(渗透压500mOsm·kg-1水,)只能短时间使用。糖醇的渗透压较低,但静脉注射的水梨醇,有20~40%都经肾脏排出,而且10%山梨醇对周围静脉血栓形成的可能性,和10%葡萄糖差不多。木糖醇能引起草酸尿和草酸盐在肾脏内的沉积,还有致癌的报道,也不能用麦芽低聚糖,因为不通过肝脏,低聚糖不能水解成葡萄糖供各个组织利用。故在需经周围静脉提供大量能量时,只能应用脂肪乳剂。如商品“Intralipid”,没有多少副作用,已广泛用于临床。
中心静脉所输液体很快能被血液衡释,不必考虑渗透压的问题,可用高达25%或50%葡萄糖浓溶液。
4.3.2 构成细胞和组织
每个细胞都有碳水化物,其含量约为2~10%,主要以糖脂、糖蛋白和蛋白多糖的形式存在。分布在细胞膜、细胞器膜、细胞浆,以及细胞间基质中。
细胞膜有两层。外层糖被膜,主要由内层伸出的糖链所组成。内层质膜,由两层脂质(磷脂和糖脂)排列而成。脂质的脂肪链是非极性的,外层的向内,内层的向外,构成脂质双层(图4-14)。糖、磷酸及其他极性基团都在质膜的两侧。有些蛋白质嵌在脂质双层内,如系糖蛋白或蛋白多糖,糖链酸及其他极性基团都在质膜的两侧。有些蛋白质嵌在脂质双层内,而紧贴在膜上。嵌在膜内的有成纤维细胞、神经胶质细胞内皮细胞上的硫酸类肝素;紧贴在膜上的有肝细胞表面的透明质酸受体和硫酸类肝素。在体温下,脂质双层是液体的。脂质分子和蛋白分子都能在质膜内移动。质膜各组分的含量依细胞而不同,肝细胞质膜含蛋白质65%、脂质30%、碳水化物近5%。
除每个细胞都有碳水化物外,糖结合物还广泛存在于各组织中。脑和神经组织中含大量糖脂,主枯分布在髓鞘上。肾上腺、胃、脾、肝、肺、胸腺、视网膜、红细胞、白细胞等都含糖脂。
消化道、呼吸道分泌的粘液中有糖蛋白。骨和腱中的类粘蛋白,血浆中的前白蛋白、α1-、α2-、β-、γ-球蛋白、凝血酶原、纤维蛋白原、运铁蛋白,激素中的甲状腺素、促甲状腺激素、促卵腺激素、促红细胞生成素,酶中的蛋白酶、核酸酶、糖苷酶、水解酶等都是糖蛋白。
蛋白多糖则存在于骨、软骨、肌腱、韧带、角膜、皮肤、血管、脐带、关节液、玻璃液中。
结缔组织的细胞间基质,主要是胶原和蛋白多糖所组成。
4.3.3 传递信息
多少年来,一直认为传递信息是蛋白质和核酸的事,碳水化物主要功用是供给热能和构成组织。近30年来,积累了大量实验证据,到1968年已公认碳水化物能携带信息,以后又有很大发展,具体表现在:
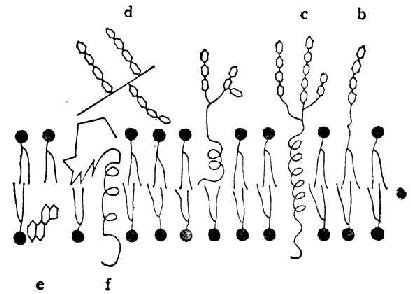
图4-14 质膜构成的示意图
а脂质双层,黑点代表极性基因,曲线代表脂肪链。
b嵌在膜内的糖脂,糖链伸在质膜外。 c嵌在膜内的糖蛋白,糖链伸在质膜外
d紧贴在膜上的蛋白多糖。 e胆固醇 f蛋白质分子
(1)作为糖蛋白和细胞被识别的标记细胞和细胞是能够相互识别的。他们可认出对方是和自己相同的,还属于另一脏器的,或者是属于另一个体的。如把肝脏、肾脏和胚胎的脑细胞放在同一个培养基里,他们会各自集中在一起。因为他们能认出伸出在对方细胞膜上的糖链是和自己相同的。又如将一个人的器官移植到另一个人身上去,会受到接受者白细胞的排斥和破坏。原因之一是他们能认出植入器官细胞膜上的糖链,和自身的不同。
在较简单的事例中,可把糖的识别作用看得更清楚。
机体中有些糖蛋白在完成一定任务后要被清除掉,细胞老化后也要处理。而正在发挥作用的糖蛋白和细胞则仍留在体内。二者的区别在完成任务或老化后他们糖链的结构起了改变。肝细胞质膜上的受体能识别这种改变了结构的糖链而结合他们,使他们进入肝细胞内,由溶酶体来使之降解。正在发挥作用的肝细胞不结合,因而不被销毁。
哺乳动物物肝脏,至少有四种能识别糖蛋白的受体:
①脱唾液酸后的糖蛋白的受体以人的免疫球蛋G为例,其低聚糖链的末端是唾液酸。唾液酸被酶解以后,末端是半乳糖。肝细胞能识别并结合末端是半乳糖的免疫球蛋白G,而不结合末端是唾液酸的免疫球蛋白G。
经这个受体清除的血清糖蛋白已知有21种,包括细胞反应的调节者促红细胞生成素、促卵泡成熟激素、绒毛膜促性腺激素、干扰素,载体蛋白如血浆铜蓝蛋白,血红素结合蛋白、维生素B12传递蛋白、皮质甾结合球蛋白、血型糖蛋白、α酸性糖蛋白、凝血酶原等。
这个受体本身也是糖蛋白,分子量41000,含糖10%。
老化的红细胞膜上的唾酸液含量,较新生的红细胞为少,能被肝细胞识别而被吞噬。新生的红细胞由于糖链末端有唾液酸,肝脏不结合,仍留在体内发挥作用。
②岩藻糖受体在肝细胞膜上,结合人乳运铁蛋白。
③磷酸甘露糖受体在肝细胞膜上,结合人乳运铁蛋白。
④甘露糖或N-乙酰氨基葡萄糖受体在枯否细胞表面。
这些受体还存在于睾丸、脾脏、肺和肾中。寿命终止的红细胞也能被脾脏识别、结合和降解。
碳水化物所担负的这种识别作用,不仅有助于保持内环境的稳定,而且还能应用于临床。如:A靶向给药,把有生物活性的分子,结合在以半乳糖为末端的低聚糖链上,可以有选择性地到达某些细胞,如肝细胞上;B诊断,去掉唾液酸后的血清糖蛋白,能灵敏地被测定,可用来评价肝硬化、肝炎、原发性肝癌中肝细胞的损伤程度。又如完整的和脱唾液酸的运铁蛋白的比值,可用来估计肝肿瘤的大小。
(2)有抗原作用有些低聚糖有抗原作用。如人奶中的游离低聚与作为载体的多肽结合后,注射入体内,能产生抗体。这种抗体能识别作为抗原的低聚糖。糖链结构稍有改变,抗体就不结合他。多糖本身就有抗原作用。
决定血型分类的是红细胞膜上糖蛋白和糖脂的低聚糖链末端的糖分子。这种糖链由14个单糖分子组成,末端4个见图4-15。A型血者最末一个单糖是N-乙酰氨基半乳糖,B型血者是半乳糖,AB型血者这两种抗原链都有,O型血都没有这两个单糖分子,链上只有13个单糖分子。

图4-15不同血型抗原的低聚糖链的末端四个单糖
肿瘤细胞上也有能起抗原作用的糖链。如NIL细胞有一种糖链,糖链末端是唾液酸。在体外把这种细胞恶性化以后,糖链末端缺少了这个唾液酸分子。把恶性化的NIL细胞给地鼠注射后,在发生肿瘤的同时,地鼠血清中产生能与缺唾液酸的糖脂产生沉淀和补体结合反应的抗体。随着从质膜上分离糖脂和糖蛋白的手段越来越精细,以及分子生物学和医学的进步,这类抗原抗体的反应,很有可能用于癌症的治疗。
(3)在细胞和细胞的粘着中发挥作用细胞和细胞可粘着在一起。粘着的方式有两种:
①直接粘着细胞表面有糖基转移酶和糖脂或糖蛋白,这个酶可与糖(如半乳糖)结合,同时把糖加于另一细胞的糖脂或糖蛋白上,使这两个细胞粘连在一起,如图4-16。正常细胞上酶和糖脂或糖蛋白的距离远,不能在独自一个细胞上结合,只能与另一个细胞相结合。肿瘤细胞二者较近,可自身结合,所以肿瘤细胞容易转移。鸡胚神经细胞、血小板、正常和恶性化的成纤维细胞已证实在其表面有糖基转移酶。β-半乳糖苷酶可破坏这种粘着作用,说明图4-16中的糖,可能是半乳糖。
②通过细胞间基质中的一种物质,与两个细胞结合已分离得到的有CSP糖蛋白,LETS糖蛋白、半乳糖蛋白α、纤粘连蛋白等。正常细胞能产生CSP和LETS;恶性化后不产生,所以细胞粘着少。如在恶性化的鸡胚成骨细胞或鼠肾细胞培养基中加入CSP或LETS,可增加他们的凝集。
图4-16 细胞粘着示意较图
e 代表酶 G 代表糖蛋白或糖脂 S 代表糖
细胞粘着失调可带来疾病。人的血小板表面有一种糖蛋白,低聚糖链末端是唾液酸。血管破裂后,血小板可通过唾液酸粘连在暴露出来的血管内皮上,起止血作用。BernardSoulier综合征患者血小板膜上这种糖蛋白含量很低,不易止血。
(4)细胞的接触抑制细胞培养时,正常细胞增加到一定的密度就不再生长。这种对生长的抑制作用,叫接触抑制。肿瘤细胞失去的接触抑制,在培养基里,密度比正常细胞高得多。这种抑制和细胞表面的糖链有很大的关系。
细胞膜的化学结构,在生长和代谢的不同阶段有所不同。在细胞生长时,一些糖链是暴露的,如肿瘤细胞和早期的胚胎细胞。正常细胞在有丝分裂时,某些糖链也是暴露的,但在有丝分裂后的短时间内,能合成一层蛋白质把这些糖链遮盖起来,使细胞互相接触后,质膜上的腺苷酸环化酶活力增加,细胞内cAMP浓度升高,把信息传到细胞核上,停止DNA的合成。肿瘤细胞能合成这层蛋白质,但边合成边被蛋白酶水解,所以不能有效地接触而推动抑制作用。胰蛋白酶能水解这层遮盖用的蛋白质,使正常的成纤维细胞失去接触抑制而不断生长。人鼻咽癌的上皮细胞和星形细胞瘤的细胞,可受二丁基cAMP(cAMP的一种衍生物,较cAMP更易进入细胞)的抑制,都是这一学说的证据。除此以外,细胞表面糖链结构的改变也是失去接触抑制的一个原因。已知的有肿瘤细胞的糖蛋白的唾液酸含量的增加,和糖脂的含量减少等。
4.3.4 起润滑作用
糖蛋白和蛋白多糖有滑润作用。关节液中有大量透明质酸,是关节活动的润滑剂。消化液中的糖蛋白,使食糜易于移动,且可包裹食糜和粪便,使肠粘膜免受机械和化学的操伤。呼吸道的糖蛋白,有防止支气管和肺泡上皮干燥、保护呼吸道免受气体和微生物侵入的作用。生殖系统的糖蛋白和蛋白多糖,有润滑和利于精子运动,保护胚胎等作用。
4.3.5 保护蛋白质不被蛋白酶消化
有些蛋白质,如酶和消化液中的糖蛋白,平常不被蛋白酶所消化,是由于分子中的糖链在保护他们。糖链结构的改变,如頜下腺分泌的糖蛋白去掉末端的唾液酸后,即被消化道的蛋白酶消化。
4.3.6 控制细胞膜的通透性
伸出在质膜外的糖链和其他极性基团,能控制水分子、无机离子和小分子的有机物的移动和进入细胞内部。如设法使糖链不能合成到蛋白上去,葡萄糖就不能进入鸡胚成纤维细胞中去,糖代谢发生障碍。
4.3.7 起蛋白质节省作用
食物中如碳水化物不足,机体不得不从蛋白质取得能量。因为能量需要量的迫切超过其他营养素。如要最大限度地把氨基酸用于蛋白质合成,在摄取必需氨基酸的同时,一定要有足够的碳水化物供应。
4.3.8 保证脂肪的充分氧化
食物中碳水化物不足,机体要利用储存的脂肪来供给能量。但机体对脂肪酸的氧化能力有一定限度。动用脂肪过多,其分解代谢的中间产物(酮体)不能完全氧化,因而引起酮病。膳食中的碳水化物可保证这种情况不会产生。
4.3.9 起解毒作用
肝中的葡萄糖醛酸能结合一些外来的化合物,以及细菌产生的毒素等,共同排出体外,起到解毒作用。
4.3.1 0作为合成生物大分子的前体
体内许多物质是利用碳水化物来合成的,如嘌呤、嘧啶、某些氨基酸、卟啉、胆固醇等。再由他们合成核酸、蛋白质及胆固醇的一些衍生物。食物中的戊糖人利用得很少。组成核酸的核糖和脱氧核糖是在体内自行合成的。
4.3.1 1膳食纤维的生理功用
人不能消化膳食纤维,但结肠内细菌的酶能使纤维素、半纤维素和果胶分解。所以大便中排出的纤维素只有食物中的20~70%,半纤维素15~45%,果胶约10%,麦麸约70%。分解产物是氢、二氧化碳、甲酸、乙酸、丙酸和丁酸等。分解时也产生能量,人能利用其中的一部分。但其数量是微不足道的。
人虽不能利用膳食纤维,但他们仍有一定的生理功用。在口腔里,增加咀嚼,刺激唾液的分泌。咀嚼时间延长,也增加胃液的分泌。对小肠的吸收也有影响。如吃相当于1004kJ(24kCal)的苹果或苹果汁,第一小时仍未恢复。因为苹果汁中的糖吸收很快,刺激了胰岛素的分泌;苹果中有果胶,使糖逐渐吸收,避免了过多的胰岛素分泌。膳食纤维还可增加胆汁的分泌,吸附胆汁酸;缩短食物残渣通过大肠的时间,增加粪的重量,减少粪的硬度。
4.4 膳食纤维和一些疾病的关系
由于人不能消化和利用膳食纤维,所以长期以来认为它们除增加肠蠕动,防止便秘外,别无他用。1956年Cleave提出发达国家中的某些疾病由于食物中除去大部分膳食纤维(以精白面粉和蔗糖作碳水化物的来源)而引起。六十年代起,在发展中国家工作的西方医生,发现某些疾病在当地居民中远较发达国家为少。1972年Painter在食物中添加麦麸治疗憩室病得到良好的效果,更引起了对膳食纤维的重视。
4.4.1 与膳食纤维有关的疾病有:
印度城市居民以精白米为主,农村是糙米。巴基斯坦城市人吃精白面,农村是全粉制品。在这两个国家里,糖尿病都是城市高于农村。
小白鼠饲料中粗纤维占5%时出现高血糖,但不见于>75%时。
用粗纤维或果胶治疗糖尿病有不少的报道。例如,13名用药治疗的糖尿病人,原来膳食中碳水化物占总能量的43%,脂肪占34%;改成碳水物占75%,脂肪占9%,并将粗纤维增加3倍。14天后用硫酰脲者全部可以停药。每天用胰岛素15~18单位的5人中4人停用。原用胰岛素40~55单位者3人可以降低剂量。15个月后随访,膳食中碳水化物占总重量的55~60%,停药的10人中7人仍不用药,血糖不高。
为澄清上述治疗效果是来自膳食纤维还是碳水化物,另一组学者将8个胰岛素的糖尿病者分为两组,碳水化物都是42%,膳食纤维分别为每天20和3g,胰岛素用量不变。10天后高纤维血糖平均1210,低纤维组平均1690mg·L-1。
高纤维组糖的吸收慢,胰岛素分泌比较平稳。所以膳食纤维对糖尿病患者有一定帮助。
4.4.2 缺血性心脏病
膳食纤维的多少和血清胆固醇浓度有一定关系。Harding和Stare报道素食者是2060,荤食者是2910mg·L-1。
膳食纤维少者,能量摄取多。胰岛素分泌增加,动脉容易硬化。胆汁酸在粪便中排出少,因而血浆胆固醇升高。这些都是缺血性心脏病的因素。给柑桔果胶3周,每天15g,血浆胆固醇降低13%;粪内脂肪增加44%,中性固醇类物质增加17%,胆汁酸增加33%。
4.4.3 胆石病
从尸检发现发达国家和发展中国家患胆石的百分率有很大差别。前者高,后者低。
胆石形成的原因是胆固醇合成过多胆汁酸合成过少。胆石病人在膳食中每天加麦麸20~40g,6周后胆汁中胆固醇含量降低。没有胆石的妇女,每天给麦麸30g,两个月后胆固醇在胆汁中的浓度也降低。
4.4.4 便秘和刺激性肠综合征
膳食纤维少后,大便量少、硬、成小块。发达与发展中国家的粪重有很大的区别,如英国平均每天重100~150g,乌干达农民470g。用X线观察小丸通过胃肠道的时间,英国是40~100h,乌干达36h。
刺激性肠综合重的症状有消化不良、食欲减退、胃灼热、恶心、腹胀、饱满感、无痛腹泻、交替的便秘和腹泻、腹痛甚至结肠铰痛等。现在认为是膳食纤维缺乏引起的肠功能改变。增加膳食纤维常有良好的效果。如每天增加麦麸30g,6周后腹痛的次数和程度、心及结肠活动都显著减少。特别是麦麸有调节肠习性的功能,每天吃30g,原来平均3.8天大便一次的可缩短到2.4天,原来1.0天一次的可延到1.72天。
4.4.5 憩室病
过去认为憩室病要用低渣膳食,现在正相反。70个患者改成高纤维膳(包括每天3~45g麦麸)后,62人中有36%症状减轻,52%症状消失。另48个患者以麦麸饼干增加膳食纤维4.5g,为期6个月到3年,73%的患者症状得到控制,33%%疼痛消失。用安慰剂作双盲法试验得到类似结果。
结肠内容物少后,肠腔狭窄,易形成闭合段,从而增加肠内的压力。同时,粪便硬和粘,需要更大的压力来排便,易得憩室病。膳食纤维增加粪的体积,能吸水,可降低硬度和粘度,有一定好处。
4.4.6 大肠癌
大肠癌的致癌物质,一般认为在粪中,可能是细菌的代谢产物如脱羟基的胆汁酸、脱氧的胆酸盐、氨、酚类等。而且肠内还常有亚硝基化合物。
流行病学调查说明大肠癌的发病率与食物中的肉类、脂肪、蛋类和总能量呈正相关,与谷类和豆类呈负相关。
大白鼠可用氧化偶氮甲烷或二甲肼引起结肠癌。在半合成饲料中如以麦麸代替蔗糖,得癌的鼠数从75%降到52%。饲料中加麦麸后,每只鼠的肿瘤数从平均6.4减少到2.7,长瘤的鼠数也从100%降到67%。
膳食纤维少后,大便量少,肠内水分少,致癌物质的浓度相对增高;粪在肠内停留时间长,细菌产生的致癌物质多,与肠粘膜时间长。在膳食纤维多的情况下刚相反。同时,纤维多后能量相对降低,而高能量膳食是容易致癌的。
4.4.7 龋齿和牙周病
龋齿、拔除和填补的牙数,发展中国家平均每人不到2枚。美国是10~25。英国3~4岁小孩平均3枚,15岁的少年8.4枚;2004人中84%有牙周病。
咀嚼增加唾液分泌,并增加其缓冲酸碱的能力,减少附着于牙上的食物,有利于防止牙周病。
4.4.8 痔和其他肛门疾病
高纤维膳食可使90%找高肛压痔疮患者的症状消失,有类似括约肌切开或肛门扩张的效果。高纤维膳食也可缓解肛裂的症状而避免手术。膳食纤维对肛门疾病有益已为许多学者所接受;低纤维膳食能否引起这类疾病则尚无定论。
4.4.9 膳食纤维对无机离子吸收的影响
膳食纤维可结合无机阳离子,减少他们以及磷的吸收。表4-15是用猪作试验的结果,对钙、镁的影响比较大。表4-6是5名男子和4名女子的结果,膳食纤维高者钙的负平衡相当严重。但有能适应这种情况,不过需6周或更长的时间。在这段时间内,维生素D摄入不足者不晒太阳者是有一定的危险的。
表4-5 纤维素对无机离子吸收的影响
| 低纤维素饲料 | 高纤维素饲料 | |||
| 摄入量(g) | 吸收率(%) | 摄入量(g) | 吸收率(%) | |
| 钠 | 5.1 | 99 | 5.6 | 98 |
| 钾 | 4.6 | 97 | 4.3 | 86 |
| 钙 | 13.5 | 74 | 13.2 | 63 |
| 磷 | 9.9 | 81 | 10.2 | 74 |
| 镁 | 1.1 | 73 | 1.4 | 59 |
| 锌 | 0.12 | 60 | 0.13 | 76 |
表4-6 白面包和黑面包对钙吸收的影响
| 白面包* | 黑面包 | |||
| 钙摄入量 | 钙平衡 | 钙摄入量 | 钙平衡 | |
| 男子 | 580.2±40.0 | -26.6±25.2 | 589.2±24.6 | -111.6±26.9 |
| 女子 | 425.8±15.3 | ± | 520.3±9.8 | -56.8±9.7 |
单位:mg·d-1
*白面包69粉,黑面包占总量能量的40~50%。
此外,膳食纤维可减少铁的吸收。铁不足者,特别是老年人和月经期的妇女,膳食纤维不宜过多
4.5 膳食中碳水化物的来源
食物中碳水化物的来源有五大类:谷物、蔬菜、水果、奶和糖。谷物中除淀粉和膳食纤维外,还有蛋白质、矿物质和维生素。薯类、豆类和植物的根和块茎都是淀粉的来源。所有蔬菜都有纤维素、蛋白质、矿物质和维生物。豆类还有脂肪。水果中有葡萄糖和蔗糖、膳食纤维、矿物质、维生素。糖是纯碳水化物,不含其他营养素,多吃后能影响食欲,降低其他营养素的摄取量。
动物性食物中只有奶能提供一定数量的碳水化物。乳糖在肠内停留时间较其他双糖长,有利于细菌的生长,某些细菌能产生维生素B12和其他B族维生素。人成年后乳糖酶逐渐消失,所以奶及奶制品会引起某些人的腹泻。
一般食物成分表上只有粗纤维的数字。表4-7列出几种食物膳食纤维含量。
表4-7 几种食物中膳食纤维的含量
(单位:每100克可食部分中的克数)
| 纤维素 | 果胶 | 木质素 | 膳食纤维总量 | |
| 麦麸 | 8.0 | 3.2 | 44.0 | |
| 白面包 | 0.7 | 痕量 | 2.7 | |
| 全粉面包 | 1.3 | 1.2 | 8.5 | |
| 苹果 | 0.5 | 0.5 | 0 | 1.4 |
| 番茄 | 0.4 | 0.3 | 0.3 | 1.4 |
| 马铃薯 | 1.0 | 0.2 | 痕量 | 3.5 |
| 卷心菜 | 0.7 | 0.7 | 0.4 | 2.8 |
| 胡萝卜 | 1.5 | 0.9 | 痕量 | 3.7 |
参考文献
1.MontreuilJ:Glycoproetins,I nComprehensive Biochemistry (Neuberger A&Van DeenenLLM),Vol 19b PartsⅡ,pp.1 ~188,Elsevier,Amsterdam,1982
2.Hakomori SJ:Ann Rev Biochem50:733-64,1981
3.Kuause MV&MahanLK:Carbohydrates,in Food,Nutrition&DietTherapy,7th ed,pp,24 ~29,Saunder,Philadeiphia,1984
4.Silk DBA:Intestinal Absorptionof Nutrients,in Surgical Nutrition(FischerJE),pp.19-25,Little,Brown,Boston,1983
5.Pike RL &BrownMLDigestion,in Nutrition,an Integrated Approach,3rd ed,pp.201-83,John Wiley&Sons,New York,1984
6.Vester JW &RudneyH:Carbohydrates,in Surgical Nutrition (Fischer JE),pp,165 ~ 212,Little,Brow Boston,1983
7.Armstrong FB:Pentose phosphatein Biochemistry,2nd,ed,pp,288 ~ 93,OxfordUniversity Press, New York,1983
8.Orten Jm&NeuhausOW:Metabolic interrelationship of tisseus,iu Human Biochemistry,10thed,pp,66382,Mosby,St.Louis,1982
9.Levenson SM&SeifterE:Starvation:metabolitc&physiologicres,in Surgical Nutrition(FischerJE),pp.423 ~ 445,Little,Brown,Boston,1983
10.Orten JM&Neuhaaus OW:HumanBiochemistry,10th ed.,p.690,Mosby,St.Louis,1982
11.LevyR1:Prevalence&etiology of cardiovascular diseases in Ceicil Textbook ofMedicine(Wyngaarden JB&Smith LH),16th ed.,pp.98 ~ 101,Saunders,Philadelphia,1982
12.Albrink MJ&UllrichIH:Effects of dietary fiber on sucrose-induced lipemia in humans,in MetabolicEffects of Utilizable Dietary Carbohydrates(Reser S),pp.221 ~ 38,Marcel Dekker,New York,1982
13.Palumbo PJ et al:Short-termsimple carbohydrate loading in patients with coronary artery disease:effectofplasma glucose,serum lipids,plasma levels of in insulin&growthhormone,metabolicrate,&blood pressure,in Metabolic Effects of UtilizableDietary Carbohydrates(Reiser S),pp.315 ~36,MarcelDekker,New York,1298
14.Jeejeehoy KN&MarlissEB:Energy supply in total parenteral nutfition,in SurgicalNutrition(FischreJE),p.647,little,Brown,Boston,1983
15.Williams ER&CahendoMA:Nutrition,Mc Graw-Hill,New York,P.62,1984
16.Pike RL&Brown ML:Theplasma membrane,in Nutrition,an Integrated,Approach,3rd ed,pp.337 ~67,John Wiley&Sons,New York,1984
17.Ashwell &Harford J:Ann RevBiochem51:531~54,1982
18.张荫昌、赫明昌等译:癌的分子生物学,195~213页,人民卫生出版社,北京,1983
19.Royal Collegeof Physicians:Medical Aspects of Dietary Fibre,pp.1 ~175,Pitman,Kent,1980