�ڶ��ڡ���֬����
֬�����еĵ��ײ��ֳ�Ϊ��֬���ף�apolipoprotein/apoprotein��Apo������֬������֬���״�л�о�����Ҫ���������ܡ�Apo���ɲ��ȶ�֬���Ľṹ�����β�Ӱ����֬���״�л�йص�ø�Ļ��ԡ���Ϊ֬������������壬����֬������ϸ������֬��������Ľ�ϼ����л���̡�
һ����֬���������ṹ�ص㼰��������
Apo����ܶ࣬һ���Ϊ5-7�࣬�䰱�������д�����Ѳ�����Apo����������ǰ�1972��Alaupovic�����������������Ӣ����ĸ˳����룬��ABC˳��ÿһ������ࡣ
��һ����֬����A��
ApoA�ɷ�ΪApoA��A��,A����ApoA���A��ֲַ���HDL�У���HDL����Ҫ��֬���ס�
1.ApoA��ApoA����ApoA������һ����ݣ��Ⱥ����HDL�з��봿���õ�ApoA��������ApoA��İ��������У�ҲԤ����������ṹ��Ҫ�㡣�˳����ApoA����243��������л���ɣ��ǵ�һ��������������Ϊ28.3ku���˼�����á�ţ��Ѽ�������ȶ����ApoA���ѷ��봿�����˺�����������ApoA��İ���ĩ��ΪAsp���Ȼ�ĩ��ΪGln��������в������װ�����������ᡣ���ȵ��۽���Ӿ֤ʵ���˺Ͷ����ApoA���Dz���һ�ģ���10�ֲ�ͬ������ݣ�������6�ֶ�̬�ԡ�
Ŀǰ��֪��ApoA��İ�����л������������������������ټ�������л������϶࣬����1��2��1��4�෴���ӶԵ���ʽ���У���Glu-Arg��Glu-Lys��Asp=lys��Asp-Arg������ˮ������л�һ�ԶԵس�����1��2��1��4�����ӶԵĸ�������˺������γ����е�˫�����������ṹ��������Ǽ�������л����еķ�ʽ����֬����һ�����ԡ�����ˮ������л���������ķǼ����棬�ɴ������ˮ�İ�����л�������������棬�ʳ�Ϊ˫����������һ�㵰���ʵĦ�-������ͬ������˫������������֬��һ��������ˮ��һ�档ApoA��˫�������ṹ������ά�������������������Ƿdz���Ҫ�ġ�
ApoA����Ҫ������HDL�У���HDL3��ApoA��ռ��֬����65������HDL2��ApoA��ռ��֬����62������CM��VLDL��LDL��Ҳ���������ڡ�Ѫ���г��֦�Ǩ���ʵ�һ�֦�-HDL������80��ΪApoA��
������������У��������֬���ײ�ά����ṹ���ȶ����������ԡ�ʵ�������������ApoA����ˮ��Һ�п����Է��غ�֬���ϡ���CNBr����ApoA���ѽ���ĸ��ĶΣ����ֽ����Ȼ�ĩ�˵��Ķο��Է�����֬��ϡ�������һ��ȷ����һ����ApoA��224-242�Σ���һ�μȿ�ά��˫�������Ľṹ���ֿ���ά�ֺ�֬�ʽ�����߱�����ˮ�ԡ���ApoA����Լ�������֬���̴�����ת��ø��LCAT���Ļ��ԡ��Ѿ�֤ʵ��ApoA����ͨ������LCAT���ٴ����̴�������ApoA���Ķ��Ķ�116-151���Ǽ������õ����ġ���ApoA�����ΪHDL��������壬��ApoA��֬�����Ժ�ת������ͭ�������γɴ���Ӹ���������������ͭ���ӡ�
���ɸκ�С���ϳɣ�Ѫ�������������Ϊ45�졣
2.ApoA��ApoA����HDL�еڶ��ֺ��������֬���ף���HDL2��ռ��֬����15������HDL3��ռ��֬����25������CM��ռ��֬����7��-10����VLDL��Ҳ������������1985�꣬ApoA���ʵİ��������У�cDNA���м��������о��Ѳ�����ApoA������������������77��������л���ɡ�ApoA���ڲ��ӻ�ԭ����SDS-PAGE�в����������17ku������Ѫ�����Զ�������ʽ���ڡ�ApoA��ĵ��������Ϊ8.7ku��ApoA��C�˰�����л�Ϊ�Ȱ��ᣬN��Ϊ������ͪ�ᣬȱ���鰱�ᡢ�����ἰɫ���ᡣApoA���ж�̬�Դ��ڡ�
ApoA�����������ǣ���ά��HDL�ṹ��ApoA���Ķ�12-31���Ķ�50-77��������֬��ϵ��������������ṹ������Ϊ���л�17-30��51-62�γɵ�˫�������ṹ����ApoA����֬�ʽ�ϵķ��ӻ������ڼ����֬ø������ˮ��CM��VLDL�е�TG��PL�����б�����ApoA�������LCAT���ԡ�
ApoA���ɸκ�С���ϳɡ���Ѫ���е�ApoA�����������Ϊ4.4�졣
3.ApoA�����ȴӴ���HDL��CM�з�����֬����A�����Ժ�֤ʵ��Ѫ����Ҳ��ApoA�����ڣ���Ҫ�ֲ����ܶȴ���1.211g/ml���֡�����ApoA����376��������л���ɡ���SDS-PAGEȷ�ϴ������ApoA��������Ϊ44-46ku���˺ʹ�����������ƣ���һ���ǵ��ף�����6����̼ˮ��������и�¶��ռ1.8����������ռ1.55����N-���������ǰ�ռ1.55������Һ��ռ1.1����ApoA���ж�̬�Դ��ڣ����������Ϊ10Сʱ��
ApoA����������Ŀǰ�в���ȫ��������Ʋ�ApoA���ڵ��̴�����ת�˹�����������Ҫ���á������ApoA�����Դٽ�LCAT�ĵ��̴�������Ӧ������Ϊ��LCAT�ļ������ApoA���ɸκ�С���ϳɣ��������ֶ�̬�͡�
������Apob��
����ʵ�������ApoB��������ˮ�ĵ����ʡ�Ŀǰ��֪��ApoB��ɷ�Ϊ�������࣬��ApoB48��ApoB100��ApoB��Ҫ�ɷ���B100�����ΪB48��������ʽ����ApoB75��ApoB41��ApoB36�Ⱦ�ΪApoB100�IJ�ͬ������
1.ApoB100ApoB�ǵ����ǵ��ף�������Ϊ51ku����Ҫ�ڸΣ�������С���ϳɡ�ApoB100��4536��������л���ɡ�����27������24���������ź��ĺ�4536��������л��ij��쵥�嵰�ס�1986��ApoB100��ȫ��������л�����˳�ṹ�Ѿ������������Ѳ��ApoB100��cDNA���У�ApoB100�����к���25��Cys�л���������11��Cys�л����зֲ���ǰ��500��������������γ����ڶ����������N�˸߶Ƚ����ɵ������νṹ��Cys�л�ͨ������������֬�ᡢӲ֬�����ϣ�ʹApoB�ι̵�������֬�ʳɷ֡�ApoB100�У���֬����ʮ�ֱ�Ҫ������ṹ��203-2506��4002-4527������л�֮�䡣�����ṹ�����ظ�����������֬��-�������Σ�����һ�ֽ��֬�ʵ���Ҫ�ṹ�Ǻ���ˮ����ˮ�����ύ�����е�������֬��-�۵��ṹ�����ֽṹ�ֲ����������������У����������ĸ������������������ָ�����������ظ�������ApoB�����еģ�ʹApoB�ܹ�����֬����������䲢ʹ֮���ܽ�ϡ�����ApoB�����Ԧ�-�����������������ˮ���Լ��ɱ�֬������Cys�л��γɵ�����ṹ����VLDL��LDL�ӷ��ڵ�����������������У�ʹ��-�����ܹ��뵥�㼫��֬�ι̵ؽ�ϣ��Ӷ�ʹ�䲻��֬�����Ӽ�ת����������������֬���ײ�֮ͬ����
ApoB100�����������У��ٺϳ�װ��ͷ��ڸ�������������VLDL������LDL�Ľṹ���ס���LDL��������壬���ɵ���LDL��Ѫ���е�������ʡ�
2.ApoB48ApoB48���������ApoB100��48����������������CM�У���������֬�����ӽ�����ApoB48��С���ϳɣ�����װCM���������֬���ס�С��ϸ������CM������ܰ�Һ����ͨ���ص����ٽ���ѪҺѭ�����ٷֲ���ëϸѪ�ܵ���Ƥϸ������Ҫ�ǹ��������֬����֯����Ƥϸ����֬��ø��ˮ��CM�и���������80��-90����ʣ�µ�֬���������ΪCM�����������͵��Σ�����֬ø��һ����л������ܹ�ʶ��ApoE�IJ���������ȡ���ò���������LDL���壬������LDL������صĵ������������ʣ�Ŀǰ�в������
��Ѫ����ApoB48��������ڽ�5-10���ӣ��ֽ��ٶȺܿ죬Ѫ���е�Ũ�Ⱥܵͣ�Լ�൱��ApoB100��0.1������SDS-PAGE��ӾȾɫ��VLDL����пɼ��������ApoB48����ʳ�ḻ��֬����ApoB48/ApoB100��ֵ�������ӡ�
������ApoC��
ApoC��Ŀǰ��֪��֬�����з�������С��һ�ࡣ���ȴ�VLDL�з����һ�ֺ���������֬�ĵͷ�������֬���ף�������Ϊ��֬����C���˺�˫��HDL�з�����ApoC������һ��ȷ��ApoC���������ͣ���Apoc��
Apoc����57��������л���ɵĵ�һ���������������Ѳ�����������װ��ᡢ�鰱����Ұ��ᡣ������Ϊ6625u����ApoC������ṹ����55����-�����ṹ����������֬��ϣ�����LCAT�ļ������
ApoC������79��������л���ɵ�һ��������������˳���Ѳ����������Ϊ9110u,�����ֶ�̬�ͣ�p��ֱ�Ϊ4.86��4.69���������װ����˿���ᣬ������ṹ�Ħ�-����Լռ23����ApoC��ɼ��������Դ��֬����֬��ø��LPL������ṹ�е�55-78λ������л���ά�����LPL�������õ���̵ı��������Ȼ���43-50λ������л�Ϊ��-�����ṹ��֬�ʽ������
ApoC����79��������л���ɵ�һ�����������ڵ�74λ�հ���л�������Һ�������ͬ���ֿɷ�ΪApoC��0��C��1��C��2�������࣬Ҳ�����̬�ԣ��ȵ��ֱ�Ϊ5.02��4.82��4.62��ApoC���������Ѳ����������Ϊ8764u��������ṹ�ڲ�ͬ״̬�£���-����Լռ22��-54�����ȡ�ApoC��Ħ�-�����ṹ��������֬��ϡ�
ApoC�����������У���ͬ��֬����ã�ά��֬���ṹ������Һ�г����������˫�����ӣ�������ɵ�������������֬������ɵĻ������ã����к�ǿ����֬��ϻ��ԡ���������֬������ã�ʹApoC��Ħ�-�����ṹ���ӣ�����֬�ĵ������������˶����ܵ����ƣ��Ӷ�Ӱ����֬������̬��Һ��̬��ת�䣬�������õĽ�����Ӷ���ϵ��֬���Ľṹ���ڶ���ø�м������ã�HDL����֬������������ʱ��ApoC��ͨ��HDL֬������ٽ���LCAT�Ĵ����ã���ApoC����Լ���LPL���伤����ƿ����ǣ�LPLͨ��������ѭ�����������ӽ�ϲ�������Ѫ����Ƥ�ϣ���LPL�Ӵ�CM��VLDLʱ��LPL��ͬ֬�������������֬�������ã����������֬�������ϣ����ڵ�ApoC����LPL�������ã��ı�LPL�Ŀռ�ṹ��������ˮ�����������
Apoc��Ҫ�ɸκϳɣ�С��Ҳ�ϳ�������
���ģ�ApoE
ApoE��һ�ָ���������ļ��Ե��ף���AppE��299��������л���ɣ�������Ϊ34.145ku����32��Arg��12��Lys��������Ѫ��CM��VLDL��������У���-VLDL�к�ApoE������VLDL��һ����ApoE��ѪҺ����ApoA���γɸ����塣�Ѳ��ApoE�ĵ�����һ���ṹ������ApoE��cDNA���У���ȷ��ApoE��3����λ�����칹���Լ�������Ⱦɫ���ϵĶ�λ��������Ͳⶨ������Һ��ApoE��62���Ħ�-������9���Ħ�-Ƭ�㡢11���Ħ�-ת�Ǻ�18�����������š�ApoE���ӿ��Ա���Ѫøˮ��ΪN-�˺�C-����������N-������1-191��Ϊ22ku�Ŀ������ף���������ȶ���C-������216-299��������Ϊ10ku�������̶Ⱥܸߣ����ȶ�������֬���Ľ������ApoE��Ҫ�ɸ���ϳɣ������������ԡ����������������ټ�����ϸ��Ҳ�ܺϳ�ApoE��
ApoE���������У�����LDL��������壬Ҳ�Ǹ�ϸ��CM������������壬����֬���״�л������أ���ApoE���ж�̬�ԣ���̬�������Ѫ֬ˮƽ����������Ӳ��������չ������ء�
���壩Apo(a)
���ڲⶨ֬���ף�a��[LP(a)]����Ⱥ�ֲ���Ϊ30����Ŀǰ���ø������ķ������ּ���������������Ⱥ�У�����Ѫ����Ȳ���ܴ�����0-1000mg/L�ķ�Χ��1987���¡����Apo(a)�Ļ������У����Ƶ������������У���ʾApo(a)�ķ��ӽṹ������øԭ��Ϊ���ơ�Apo(a)����һ����ˮ�ź����У�37��Kringle-4������1��Kringle-5��1���ȵ���ø��������36��Kringle-4����һ������δ����װ��ᣬ�Ʋ�˴�������Apo(a)�Զ������ApoB��ϵIJ�λ�����ȵ���ø������ˮ��Apo(a)���֣�Apo(a)�е�Krtingle-4��75��-85%�İ�����������øԭ�ĵ�391-472��������ͬ���й�ͬ�Ŀ�ԭ�أ����߱����н��淴Ӧ������øԭ��PG����һ��˿���ᵰ��øԭ������791������л����ṹ�к���5���������װ���ġ�Kringle�����ṹ����Kringle1-5����Kringle-5�ĺ���Ϊһ˿���ᵰ��ø����PG��Apo(a)�ṹ���ƣ���ͼ4-3��ʾ��
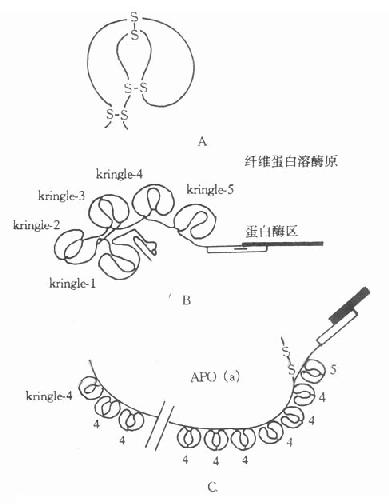
ͼ4��3 ��֬���ף�a���ṹʾ��ͼ
A:KringleB����ά������øԭC��Apo��a��
Kringle�ṹ�����Զ������ɵ������νṹ������78-82��������л����������е���д��ʽ����һ�ֵ������������������1���6����2���4����3���5���װ����ϣ�������������������ֽṹҲ������ǰ��Ѫø����ø������ø������øԭ�������t-PA��������С�����Apo(a)�����е�Kringle-4��Ŀ����15-27֮��仯���Ӷ�����Apo(a)�ж��ֲ�ͬ���칹�塣
Apo(a)�ṹ����һ����ø�����Ʋ��书�ܿ�����һ��ø���ڷ������൱��PG����ø��˿���ᱻ��������棬��ʹ��ɥʧø�Ĺ��ܡ�����Kringle�ṹ��PG���ƣ��Ʋ�Apo(a)���ܽ�ϵ���PG�������ά���������Ĵ�����ϣ��ټ���LP(a)����Я���ĵ��̴���ϵ�Ѫ�����˲�λ������������ٽ���������Ӳ���γɣ�Ҳ�谭Ѫ������Ѫ����ܽ⡣
������֬���Ļ���ṹ������
��һ�������̬�Ը���
�������ﶼ��ͨ����ֳ�����Ӵ����Ӵ����״�֮�䣬��������̬������������ܵ��ص��϶������ƣ����������Ϊ�Ŵ���heredity�������ǣ��״����Ӵ�֮�䣬�Ӵ��ĸ�����֮�䲻����ȫ��ͬ���ܻ��������죬��������б��죨variation�����Ŵ��ͱ������������������Ŵ��ͱ���������Ƕ��������ӵģ�����Ϊ��ˣ��ŵ��������Ķ��ֶ����ԣ������������е��Ŵ���״��Ϊ���ͻ�����ͣ�phernotype���������������е��������ɷֳ�Ϊ�����ͣ�genotype���������ǻ������뻷����������õĽ�����Ŵ�����������ȶ��ģ��������ǿɱ�ģ��Ŵ����ʵı仯�Լ�������������͵ĸı䣬��Ϊͻ�䣨mutation�����Ŵ�����ͻ�����Ⱦɫ�����ͻ���ͻ�䡣����ͻ����Ⱦɫ����ijһ���Ϸ�����ѧ�ı䣬�����ֳ�Ϊ��ͻ�䣨pointmutation��������ṹ���Ŵ����͵��о��������˽�֬���״�лȱ��֢�ķ�������ѧ�����������Ŵ�ѧ������reversegeneticapproach����ʹ���п����ڵ�����ˮƽϵͳ�ط����ṹ���ܵĹ�ϵ�����Ѳ���һ���ض���cDNA̽��ӻ����Ŀ���ɸѡ����Ҫ�Ļ������cDNA��¡���ⶨ����������У�Ȼ��Ӻ����������ƶϵ����ʰ��������С�Ŀǰ���ѷ���������붯������Ӳ���йص�֬����cDNA��¡�������䵰����һ���ṹ�İ���������˳��ͻ���ĺ�����˳���������Ѳ�����ApoA��A����E��B��C���(a)���������칹�壬Ҳ����˵�����Ÿ��ֲ�ͬ�ı��ͻ�����ͣ����ɷֱ�ӵ�����ˮƽ�ͺ���ˮƽ���з��͡��ֱַ���ܼ�����Ҫ��֬���Ļ���ṹ��
��������֬������ṹ�ص�
��Ѫ������֬���Ľṹ�����ܣ�������ʮ��������о������˽�ý�Ϊ���������֬���Ļ����cDNA���ѵõ������ȷ�����������˳��Ҳ�����˲ⶨ����ApoA����B��(a)�⣬���ǵĹ�ͬ�ص��Ǻ��������ں��ӣ�intron�����ĸ������ӣ�exon�������ں��Ӳ��������ӵ�λ�ô�����ͬ�������ϰ����������ܵIJ�ͬ��������Էָ�����һ���ں��Ӱ�5��-ĩ�˵ķǷ������ͷ������ֿ����ڶ����ں��Ӱ��ź��ı��루singnalpeptide�����ܵ��ױ������ֿ����������ں������ԭ�ı������ͳ����ı������ֿ�����Щ����ĵ�һ�������������ӵĺ���������Ҳ����������ĸ������Ӻ�����������ͬ�����¸�����֬�����Ȳ�ͬ������������Ƕȿ��ǣ�������֬������ṹ�����ԣ���ʾ������Դ��һ����ͬ�����ȣ���ApoC�����ApoA����������֬������ṹ��ͬ����ֻ�������������ӡ���֬������ṹ����һ�ص��Ǽ���������Ӻܽ�����λ��ͬһȾɫ���һ��λ���ϻ����ʽ�������״̬����ApoA��C���A������λ�ڵ�11��Ⱦɫ�峤��2�����γ�һ��Լ15kb�Ļ���ء�����һ�����������Ļ������ApoE��C���C�����ͬλ�ڵ�19��Ⱦɫ�峤��3������ͼ4-4��
ApoA-�����λ�ڵ�1��Ⱦɫ�峤��2����ApoB����λ�ڵ�2��Ⱦɫ��̱�2����Apo(a)����λ�ڵ�6��Ⱦɫ�峤��2����
��������֬������ṹ
1.ApoA��ApoA�����1863bp�����������ں��ӣ���һ���ں���λ��5��˷Ƿ��������ڶ����ں���λ�ڷ�������A��ǰ�����ڣ��������ں��Ӳ��뷭�����A���43������л�����ApoA��������ĸ������ӣ��ֲ���ApoA�����IJ�ͬ����ApoA�������ApoC��A�����������ɴأ�C�������У�ת¼������A���A�������෴��λ��A���C�����ͬ3������DNA���У����ܲ����A������ת¼���ء�
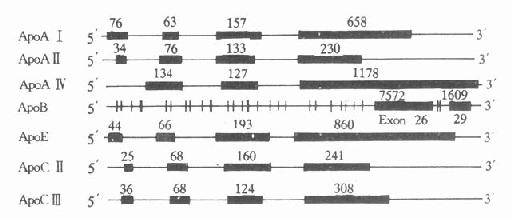
ͼ4-4 ����֬����A��A��A����B100��
C��C���E����ṹʾ��ͼ
���ߴ��������ӣ�����֮���ϸ�ߴ����ں��ӣ�������Ե����
�����öκ�������Ŀ
2.ApoBApoB��λ��2��Ⱦɫ��P23��Pter�������ɷǷ���������������TAA��ֹ�����Ӻ�һ��3��˵ķǷ�������ɡ�ApoB100����ȫ��43kb����29�������Ӻ�28���ں��Ӽ�ͼ4-4�����е�26�͵�29�����������ر��ֱ���7552��1905bp��������2��̣���39bp����211-249�����ں������Ե�27��Ϊ��̣�107bp������Ⱥ��������14�ֲ�ͬ��3��˸߱����λ��������75������Ⱥ�ڴ����Dz���һ�ġ�
ApoB48��ApoB100�����ڽṹ���й��⣬ApoB48���γɻ���Ŀǰ������ȫһ�µĿ�������Ҫ��Ϊ�кϳ�ApoB48�Ļ�����ڡ�1987�걻����ApoB48����ApoB100ͨ��һ���µĻ����漰��mRNA�ı༭�������ġ��ڲⶨ����С�����������ApoBcDNA������ʱ���֣�С��ApoBcDNA�ĵ�6666��������ΪT�����Ӹη����ApoBcDNA��¡�ڴ�λ��ΪC����T�滻C��6666������һ��ֹ���루TAA����TAA�滻CAA����ʹApoB100��2153λ������ӦΪGln��ԤʾѪ���д��ڵ�ApoB48Ӧ���൱��ApoB100��2153����ĩ��ΪGln����һԤ������õ�ʵ��֤ʵ�������ֺ�������6666���滻C��Tֻ������С����mRNA�ϣ�����������С�������飨genomic��DNA�ϣ��������ת¼�Ժ��һ�������γɵı༭С��mRNA�Ľ����
3.ApoE��ApoE����λ��19��Ⱦɫ�峤��3���������ĸ������Ӻ������ں��ӡ�
1975�����ȹ۲쵽ApoE�Ķ�̬�ԣ����õȵ�۽���Ӿ��SDS-PAGE����ȷ��ApoE�Ķ�̬�ԡ�ʵ�������ApoE�������칹�壨isoform����E2��E3��E4���е���ֻ����һ����Ҫ�칹�弴�����ӣ��е��˿ɺ�������Ҫ�칹��Ϊ�Ӻ��ӡ��ɴ˿ɼ�����Ⱥ�п������ֲ�ͬ�ı��֡�����ApoE�������ApoE����ģ����Ϊ��ApoE�ĺϳ�����λ��һ������λ���ϵ�������λ���������ƣ���E2��E3��E4��ÿһ����λ�����Ӧ��һ����Ҫ�칹�壬�������ִ����ӣ�E2/2��E3/3��E4/4���������Ӻ��ӣ�E2/3��E2/4��E3/4�������ֳ������ͣ����⣬���м��ټ����칹�塣һ����Ϊ����Ҫ�칹��������Ҫ�칹�巭�����Һ���ǻ����κ�ת�������ApoE3/3���ֳ�Ұ���͡�ApoE�Ļ������е�112λ��158λ���ְ�����л��������ᣨArg���Ͱ��װ��ᣨCys���Ľ����������칹������ࡣApoE4��������λ���϶���Arg��E2����Cys��112��158λ��Arg��ΪApoE3�칹�塣��Ȼ��Ⱥ�У�����Ƶ�ʣ�3���ֲ���ߣ�ApoE3/3���ͷֲ�Լ70������ͼ4-5��
4.ApoC��ApoC�������3347bp������4�������Ӻ�3���ں��ӡ�ApoC����Ȼ�ĩ�˰����������Ǽ���֬����֬��ø�Ļ��Թ�������ApoC�������3133bp����4�������Ӻ�3���ں��ӡ�

ͼ4-5 ��ApoE������Ҫ�칹��İ�����л�����������ĸı�λ��
5.Apo(a)����cDNA̽�����Ⱦɫ�嶨λ�о�ʱ���֣�Apo(a)�Ļ���λ�����˵�6��Ⱦɫ�峤��2��6-7���䣬��Ѫ����øԭ��PLG���Ļ���λ���в����ص����ⶨPLG������Ϊ525kb����18���ں�����19����������ɣ�5��Kringle�ṹ�ɸ������������ӱ��롣Apo(a)cDNA����������Apo(a)��PLG�Ļ����кܶ�����֮����
ͨ����ϵ�о���Ŀǰ�ѷ���Apo(a)����λ����������26����λ�������̬���йء���Щ��λ�������ٱ�����34��Apo(a)�칹�塣